20.01.2022
Nemijelinizirana i mijelinizirana nervna vlakna. Koncept "nervnih vlakana"
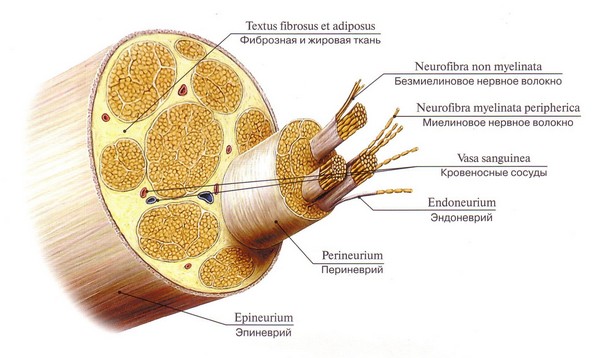
Nervno vlakno je izduženi proces neurona prekriven lemocitima i mijelinskom ili nemijelinskom ovojnicom. Njegova glavna funkcija je provođenje nervnih impulsa. U perifernom i centralnom nervnom sistemu prevladavaju kašasta (mijelinizirana) nervna vlakna koja inerviraju skeletne mišiće; Vlakna koja nemaju omotač nazivaju se goli aksijalni cilindri.
Nervno vlakno se zasniva na procesu neurona, koji formira neku vrstu osovine. Izvana je okružena mijelinskom ovojnicom s biomolekularnom lipidnom bazom, koja se sastoji od velikog broja zavoja mezaksona, koji se spiralno omotava oko neuronske ose. Tako dolazi do mijelinizacije nervnih vlakana.
Mijelinska nervna vlakna perifernog sistema dodatno su prekrivena odozgo pomoćnim Schwannovim ćelijama koje podržavaju akson i hrane tijelo neurona. Površina kašaste membrane ima intervale - Ranvierove intercepte, na tim mjestima aksijalni cilindar je pričvršćen za vanjsku Schwannovu membranu.
Sloj mijelina nema svojstva električne provodljivosti, oni imaju presjeke. Ekscitacija se javlja u Ranvierovom intervalu najbližem mjestu izloženosti vanjskom stimulansu. Impuls se prenosi naglo, od jednog presretanja do drugog, što omogućava veliku brzinu širenja impulsa.
Mijelinska nervna vlakna regulišu metabolizam u mišićnom tkivu, imaju visoku otpornost na bioelektričnu struju.
Ranvierove praznine stvaraju i pojačavaju impulse. Vlakna centralnog nervnog sistema nemaju Schwannovu membranu, ovu funkciju obavlja oligodendroglija.
Amijelinizirana tkiva imaju nekoliko aksijalnih cilindara, nemaju mijelinski sloj i presjeke, odozgo su prekrivena Schwannovim stanicama, između njih i cilindara nastaju prorezi. Vlakna imaju lošu izolaciju, omogućavaju širenje impulsa od jednog procesa neurona do drugog, kontakt sa okolinom u cijelom, brzina provođenja impulsa je mnogo manja nego kod kašastih vlakana, dok tijelu treba više energije.
Velika nervna debla nastaju od kašastih i nemesnatih procesa neurona, koji se, pak, granaju u manje snopove i završavaju nervnim završecima (receptor, motor, sinapse).
Nervni završeci su krajevi mijeliniziranih i nemijeliniziranih nervnih vlakana, koji formiraju interneuronske kontakte, receptorske i motoričke završetke.
Principi klasifikacije

Različite vrste nervnih vlakana imaju nejednaku brzinu provođenja ekscitatornih impulsa, što zavisi od njihovog prečnika, trajanja akcionog potencijala i stepena mijelinizacije. Postoji direktno proporcionalna veza između brzine i prečnika vlakana.
Strukturno-funkcionalna metoda za klasifikaciju Erlanger-Gasser nervnih vlakana prema brzini provođenja nervnog impulsa:
- Grupa A mijelinizirana nervna vlakna: α, β, Υ i δ. Najveći promjer i najdeblja ljuska su tkiva α - 20 mikrona, imaju dobru brzinu provođenja impulsa - 120 m / s. Ova tkiva inerviraju izvor ekscitacije od stuba kičmene moždine do receptora skeletnih mišića, tetiva, i odgovorna su za taktilne senzacije.
Preostale vrste vlakana imaju manji prečnik (12 mikrona), brzinu impulsa. Ova tkiva prenose signale iz unutrašnjih organa, izvora boli u CNS-u.
- Mijelinska vlakna grupe B pripadaju. Ukupna brzina provođenja impulsa je 14 m/s, akcioni potencijal je 2 puta veći od vlakana grupe A. Mijelinski omotač je slabo izražen.
- Nemijelinizirana vlakna grupe C imaju vrlo mali prečnik (0,5 mikrona) i brzinu ekscitacije (6 m/s). Ova tkiva inerviraju simpatički nervni sistem. U ovu grupu spadaju i vlakna koja provode impulse iz centara bola, hladnoće, toplote i pritiska.
Procesi neurona se dijele na aferentne i eferentne. Prvi tip osigurava prijenos impulsa od tkivnih receptora do centralnog nervnog sistema. Drugi tip prenosi ekscitaciju od centralnog nervnog sistema do tkivnih receptora.
Funkcionalna klasifikacija nervnih vlakana aferentnog tipa prema Lloyd-Huntu:
Demijenilizacija
Proces demijelinizacije nervnih vlakana je patološko oštećenje mijelinske ovojnice, što uzrokuje disfunkciju tkiva. Patologiju uzrokuju upalni procesi, metabolički poremećaji, neuroinfekcija, intoksikacija ili ishemija tkiva. Mijelin se zamjenjuje fibroznim plakovima, što dovodi do poremećenog provođenja impulsa.
Prvi tip demijelinizacije je mijelinopatija uzrokovana autoimunim reakcijama organizma, Canavan bolest, Guillain-Barréov sindrom, Charcot-Marie-Tooth amiotrofija.
Drugi tip je mielinoklastija. Patologiju karakterizira nasljedna predispozicija za uništavanje mijelinske ovojnice (Binswangerova bolest).
Demijelinizirajuće bolesti

Bolesti koje dovode do razaranja mijelinske ovojnice najčešće su autoimune prirode, drugi uzrok može biti liječenje antipsihoticima ili nasljedna predispozicija. Uništavanje lipidnog sloja uzrokuje smanjenje brzine provođenja podražajnih impulsa.
Bolesti se dijele na one koje zahvaćaju centralni nervni sistem i patologije koje oštećuju perifernu mrežu. Bolesti koje utiču na rad centralnog nervnog sistema:
- Mijelopatija kičmene moždine nastaje kao rezultat kompresije mijelinskih vlakana intervertebralnim kilama, tumorima, fragmentima kostiju, nakon. Kod pacijenata se smanjuje osjetljivost i snaga mišića u zahvaćenom području, javlja se pareza ruku ili nogu, narušava se rad crijeva i mokraćnog sistema, razvija se atrofija mišića donjih ekstremiteta.
- Leukodistrofija mozga uzrokuje oštećenje bijele tvari. Pacijenti imaju poremećenu koordinaciju pokreta, ne mogu održati ravnotežu. Razvija se mišićna slabost, pojavljuju se nevoljne konvulzije, nervni tik. Postepeno pogoršanje pamćenja, intelektualnih sposobnosti, vida i sluha. U kasnijim fazama javlja se sljepoća, gluvoća, potpuna paraliza i poteškoće u gutanju hrane.
- Malofokalna leukoencefalopatija mozga najčešće pogađa muškarce starije od 60 godina. Glavni razlozi su arterijska hipertenzija i nasljedna predispozicija. Pacijenti imaju oštećenje pamćenja i pažnje, postoji letargija, teškoće u govoru. Hod se usporava, koordinacija pokreta je poremećena, pojavljuje se urinarna inkontinencija, pacijentu je teško gutati hranu.
- Sindrom osmotske demijelinizacije karakterizira raspad mijelinskih ovojnica u moždanim tkivima. Bolesnici imaju poremećaj govornog aparata, stalni osjećaj pospanosti, depresije ili razdražljivosti, mutizam, pareze svih udova. U ranim stadijumima bolesti, proces demijelinizacije je reverzibilan.
- Multipla skleroza se manifestuje utrnulošću jednog ili dva uda, delimičnim ili potpunim gubitkom vida, bolom pri pomeranju očiju, vrtoglavicom, umorom, tremorom udova, poremećenom koordinacijom pokreta, trnjenjem u različitim delovima tela.
- Devićeva bolest je upalna autoimuna bolest koja zahvaća optički nerv i kičmenu moždinu. Simptomi uključuju različite stepene oštećenja vida, do sljepoće, parapareze, tetrapareze, poremećaja funkcionisanja karličnih organa.
Simptomi bolesti zavise od područja oštećenja mijelinskih vlakana. Možete identificirati proces demijelinizacije pomoću kompjuterske tomografije, terapije magnetnom rezonancom. Znakovi oštećenja perifernog nervnog sistema nalaze se na elektromiografiji.
Mijelinizacija aferentnih vidnih puteva počinje u lateralnom genikulativnom tijelu u petom mjesecu gestacije i završava se rođenjem na rebrastoj ploči. Oligodendrociti odgovorni za mijelinizaciju CNS vlakana normalno su odsutni u retini. Histološke studije su potvrdile prisustvo navodnih oligodendrocita i mijelina u područjima mijeliniziranih nervnih vlakana i njihovo odsustvo izvan ovih područja.
At autopsija mijelinskih vlakana retine se otkrivaju u približno 1% očiju i u 0,3-0,6% oftalmoloških pacijenata tokom rutinskog pregleda.
mijelinizirana nervna vlakna obično se pojavljuju kao uzdužno prugaste fascikule na gornjem i donjem polu diska. Istovremeno, zbog istaknutosti izmijenjenih dijelova diska i preklapanja ivica diska i krvnih žila ispod njih vlaknima, anomalija može simulirati edem optičkog diska.
Distalno se formiraju vlakna nepravilan oblik lepeze. Unutar zone mijelinizacije ponekad su vidljive male fisure ili područja normalnog fundusa. U 17-20% slučajeva otkrivaju se bilateralna mijelinizirana nervna vlakna. U 19% slučajeva ne dolaze u kontakt sa glavom optičkog živca. Ponekad se na periferiji retine na nazalnoj strani ONH otkrivaju izolirani snopovi mijeliniziranih nervnih vlakana.
Patogeneza mijelinskih vlakana ostaje nejasno, ali životinje sa slabom ili odsutnom laminom cribrosa obično imaju duboku fiziološku ekskavaciju i ekstenzivnu mijelinizaciju retinalnih nervnih vlakana, dok životinje sa dobro razvijenom kribriformnom laminom pokazuju potpuno ravan optički disk (OND) i nedostatak retinalne mijelinizacije, što sugerira nekoliko mogućih mehanizama patogeneze:
1. Prisutnost defekta na kribriformnoj ploči, koji omogućava oligodendrocitima da prodru u retinu i proizvode mijelin.
2. Moguće je da je broj aksona mali u odnosu na veličinu skleralnog kanala, te da ima dovoljno prostora za nastavak mijelinizacije u očnoj šupljini. U očima s periferno izoliranim snopovima mijeliniziranih nervnih vlakana, poremećaj vremena lamina cribrosa omogućava oligodendrocitima da uđu u retinu i migriraju u sloj nervnih vlakana dok ne dostignu područje relativno niske gustoće nervnih vlakana gdje se mijelinizacija nastavlja.
3. Kasni razvoj kribriformne ploče može stvoriti uslove za migraciju oligodendrocita u tkiva oka.
Ekstenzivna jednostrana (ili, rijetko, bilateralna) mijelinizacija nervnih vlakana može biti praćena visokom miopijom i refraktivnom ambliopijom. Kod ovih pacijenata, mijelin pokriva većinu, ako ne i sav obim diska. U makularnoj regiji (iako bez mijelina) obično se otkrivaju i anomalije - slabljenje refleksa ili disperzija pigmenta. Stanje makularnog područja je vjerovatno najtačniji prediktor efikasnosti okluzivne terapije.
mijelinizirana nervna vlakna može biti praćen Gorlinovim sindromom (višestruki nevusi bazalnih ćelija) i autosomno dominantnom vitreoretinopatijom, praćenom kongenitalnim oštećenjem vida, bilateralnom ekstenzivnom mijelinizacijom sloja retinalnih nervnih vlakana, teškom degeneracijom staklastog tijela, visokom kratkovidnošću, noćnom distrofijom retinalne sljepoće, sljepoćom mrežnice.
mijelinizirana nervna vlakna mogu se naslijediti na autosomno dominantan način. Opisani su izolirani slučajevi mijeliniziranih nervnih vlakana povezanih s abnormalnim dugim optičkim živcem (oksicefalijom), defektima lamine cribrosa (kosi optički disk), disgenezom prednjeg segmenta i neurofibromatozom tipa 2. Iako mijelinizirana nervna vlakna mogu koegzistirati s neurofibromatozom, mnogi autori ovu kombinaciju smatraju upitnom.
Povremeno zone mijelinizacija nervnih vlakana može se javiti nakon djetinjstva, pa čak i u odraslom dobu. U takvim slučajevima, trauma oka (tupa trauma oka u jednom slučaju i fenestracija ovojnice optičkog živca u drugom) je vjerovatno uobičajeni uzrok ove anomalije.
Moguće je da takva šteta postoji defekt na pločici dovoljan za prodiranje oligodendrocita u retinu. Mijelinska nervna vlakna mogu nestati kada su aksoni oštećeni.
Jednostrana miopija visokog stepena sa mijelinizovanim nervnim vlaknima. mijelinizirana nervna vlakna.
mijelinizirana nervna vlakna. A. Lagani stepen. B. Teški stepen.
Procesi neurona su gotovo uvijek prekriveni omotačem (mijelin). Izuzetak su slobodni završeci nekih procesa. Proces zajedno sa omotačem naziva se "nervno vlakno".
Nervno vlakno se sastoji od: osovinski cilindar- izrastanje nervne ćelije: akson ili dendrit
Glijalni omotač, koji okružuje aksijalni cilindar u obliku spojnice. U CNS-u ga formiraju oligodendroglija, a u PNS-u Schwannove ćelije (neurolemociti su vrsta oligodendroglije).
Nervna vlakna se dijele na nemijelinizirana i mijelinizirana (imaju mijelinsku ovojnicu).
Nemijelinizirana nervna vlakna su dio autonomnog nervnog sistema i predstavljena su aksonima efektorskih neurona. Ima ih i u centralnom nervnom sistemu, ali u manjim količinama.
Građa: U centru je jezgro oligodendrocita (lemocita), a duž periferije 10-20 aksijalnih cilindara prodire u njegovu citoplazmu. Takva nervna vlakna se nazivaju i "vlakna tipa kabla". Kada je aksijalni cilindar uronjen u citoplazmu oligodendrocita, dijelovi plazmoleme potonjeg se približavaju jedni drugima i formira se mezenterij - "mesakson" ili dvostruka membrana. Sa površine, nervno vlakno je prekriveno bazalnom membranom.
Mijelinska nervna vlakna su dio CNS-a, somatskih odjela PNS-a i preganglionskih odjela autonomnog nervnog sistema. Mogu sadržavati i aksone i dendrite neurona.
Struktura: Aksijalni cilindar je uvijek 1, smješten u sredini. Školjka ima 2 sloja: unutrašnji (mijelin) i vanjski (neurolema), predstavljene jezgrom i citoplazmom Schwannove ćelije. Vani se nalazi bazalna membrana. Mijelinski sloj je nekoliko slojeva membrane oligodendrocita (lemocita). Membrana je koncentrično uvijena oko aksijalnog cilindra. U stvari, ovo je veoma izdužen mezakson. Mesaksoni formiraju jezične citoplazmatske procese.
Proces mijelinizacije je formiranje mijelinske ovojnice. Javlja se u kasnim fazama embriogeneze i u prvim mjesecima nakon rođenja.
Vrijedi napomenuti da postoje karakteristike mijelinizacije u CNS-u: 1 oligodendrocit formira mijelinsku ovojnicu oko nekoliko aksijalnih cilindara (uz pomoć nekoliko procesa koji se rotiraju). Nema bazalne membrane.
Struktura mijelinskog vlakna.
Mijelin se redovno prekida na Ranvierovim čvorovima. Udaljenost između presjeka je 0,3 - 1,5 nm. U području presretanja vrši se trofizam aksijalnog cilindra. Mijelin ima zareze na svojoj površini. Ovi dijelovi rezanja mijelina povećavaju fleksibilnost nervnog vlakna i predstavljaju "rezervu" za istezanje. U CNS-u nema zareza.
Mijelin je obojen bojama za lipide: Sudan, Osmička kiselina.
Mijelinske funkcije:
Povećanje brzine provođenja nervnog impulsa. Vlakna bez mijelina imaju brzinu od 1-2 m/s, dok mijelinizirana vlakna imaju brzinu od 5-120 m/s.
Na-kanali su koncentrirani u području presjeka, gdje nastaju bioelektrične struje. Skaču s jednog presretanja na drugo. Ovo je saltaciona provodljivost, odnosno provođenje impulsa u skokovima.
Mijelin je izolator koji ograničava ulazak struja koje se šire okolo.
Razlika u strukturi mijeliniziranih i nemijeliniziranih vlakana.
nemijelinizirana vlakna mijelinizirana vlakna
Nekoliko osovinskih cilindara 1 osovinski cilindar
Aksijalni cilindri - aksoni
Jezgro oligodendrocita je u centru Jezgro i citoplazma oligodendrocita su na periferiji vlakna
Mesaksoni su kratki Mesaksoni se više puta uvijaju oko aksijalnog cilindra, formira se mijelinski omotač
Na- kanali duž cijele dužine aksijalnog cilindra Na- kanali samo u Ranvierovim čvorovima
Struktura perifernog živca.
Nerv se sastoji od mijeliniziranih i nemijeliniziranih vlakana grupiranih u snopove. Sadrži i aferentna i eferentna vlakna.
Mehanizmi provođenja nervnih impulsa.
Sinapse su posebne međućelijske veze koje se koriste za prijenos signala iz jedne ćelije u drugu.
Kontaktne oblasti neurona su veoma blizu jedna drugoj. Ali ipak, između njih često ostaje sinaptički jaz koji ih razdvaja. Širina sinaptičke pukotine je reda veličine nekoliko desetina nanometara.
Da bi neutroni uspješno funkcionirali, potrebno je osigurati njihovu izolaciju jedan od drugog, a interakciju između njih osiguravaju sinapse.
Sinapse djeluju kao pojačivači nervnih signala duž njihovog puta. Efekat se postiže činjenicom da jedan električni impuls relativno male snage oslobađa stotine hiljada molekula medijatora, koji su prethodno bili sadržani u mnogim sinaptičkim vezikulama. Mnoštvo molekula medijatora sinhrono djeluje na malo područje kontroliranog neurona, gdje su koncentrirani postsinaptički receptori - specijalizirani proteini koji pretvaraju signal sada iz kemijskog oblika u električni.
Trenutno su poznate glavne faze procesa oslobađanja medijatora. Nervni impuls, odnosno električni signal, nastaje u neuronu, širi se duž njegovih procesa i stiže do nervnih završetaka. Njegova transformacija u hemijski oblik počinje otvaranjem kalcijevih jonskih kanala u presinaptičkoj membrani, čije stanje kontroliše električno polje membrane. Sada joni kalcijuma preuzimaju ulogu nosioca signala. Ulaze kroz otvorene kanale unutar nervnog završetka. Naglo povećana koncentracija kalcijevih jona u blizini membrane za kratko vrijeme aktivira molekularni stroj za oslobađanje medijatora: sinaptičke vezikule odlaze do mjesta njihovog naknadnog spajanja s vanjskom membranom i, konačno, izbacuju svoj sadržaj u prostor sinaptičke pukotine. .
Sinaptička transmisija se odvija nizom dva prostorno odvojena procesa: presinaptičkog s jedne strane sinaptičkog jaza i postsinaptičkog s druge (slika 3). Završeci procesa kontrolnog neurona, povinujući se električnim signalima koji su im došli, oslobađaju posebnu posredničku tvar (medijator) u prostor sinaptičke pukotine. Molekuli medijatora prilično brzo difundiraju kroz sinaptičku pukotinu i pobuđuju električni signal odgovora u kontrolisanoj ćeliji (drugi neuron, mišićno vlakno, neke ćelije unutrašnjih organa). Desetak različitih niskomolekularnih supstanci djeluje kao posrednik:
acetilholin (estar amino alkohola holina i sirćetne kiseline); glutamat (anjon glutaminske kiseline); GABA (gama-aminobutirna kiselina); serotonin (derivat aminokiseline triptofan); adenozin itd.
Prethodno ih presinaptički neuron sintetiše iz dostupnih i relativno jeftinih sirovina i čuvaju do upotrebe u sinaptičkim vezikulama, gde su, kao u kontejnerima, zatvoreni identični delovi medijatora (nekoliko hiljada molekula u jednoj vezikuli).
sinapsni dijagram
Iznad - dio nervnog završetka, ograničen presinaptičkom membranom, u koji su ugrađeni presinaptički receptori; sinaptičke vezikule unutar nervnog završetka ispunjene su medijatorom i u različitim su stupnjevima spremnosti za njegovo oslobađanje; membrane vezikula i presinaptička membrana sadrže presinaptičke proteine. Ispod - dio kontrolirane ćelije u čiju su postsinaptičku membranu ugrađeni postsinaptički receptori.
Sinapse su pogodan objekat za regulisanje tokova informacija. Nivo pojačanja signala tokom njegovog prijenosa kroz sinapsu može se lako povećati ili smanjiti promjenom količine oslobođenog medijatora, sve do potpune zabrane prijenosa informacija. Teoretski, ovo se može postići ciljanjem na bilo koju fazu oslobađanja posrednika.
Mijelinska vlakna. Dio nervnih vlakana tokom embriogeneze prolazi kroz mijelinizaciju: lemociti (Schwannove ćelije) prvo dodiruju akson, a zatim ga obavijaju. Membrana lemocita obavija se oko aksona poput rolne, formirajući višeslojnu spiralu (mijelinsku ovojnicu). Mijelinska ovojnica nije kontinuirana - duž cijele dužine nervnog vlakna na jednakoj udaljenosti jedan od drugog, u njemu su mali prekidi (presreće Ranvier). U području presjeka, akson je lišen mijelinske ovojnice.
nemijelinizirana vlakna. Mijelinizacija ostalih vlakana završava se u ranim fazama embrionalnog razvoja. Jedan ili više aksona je uronjeno u lemocit; potpuno ili djelomično ih okružuje, ali ne formira višeslojnu mijelinsku ovojnicu.
Mehanizam provođenja ekscitacije duž nemijeliniziranih nervnih vlakana
U mirovanju, cijela unutrašnja površina membrane nervnog vlakna nosi negativan naboj, a vanjska strana membrane je pozitivna. Električna struja između unutrašnje i vanjske strane membrane ne teče, jer. lipidna membrana ima visoku električnu otpornost.Tokom razvoja akcionog potencijala u pobuđenom području membrane dolazi do reverzije naelektrisanja. Na granici pobuđenog i nepobuđenog područja počinje teći električna struja. Električna struja iritira najbliži dio membrane i dovodi ga u stanje ekscitacije, dok se prethodno pobuđeni dijelovi vraćaju u stanje mirovanja. Tako talas ekscitacije pokriva sve nove delove membrane nervnih vlakana.
Mehanizam provođenja ekscitacije duž mijelinskih nervnih vlakana
U mijeliniziranom nervnom vlaknu, dijelovi membrane prekriveni mijelinskom ovojnicom nisu ekscitabilni; ekscitacija se može javiti samo u područjima membrane koja se nalaze u području Ranvierovih čvorova.Razvojom AP u jednom od Ranvierovih čvorova, naboj membrane je obrnut. Između elektronegativnog i elektropozitivnog dijela membrane nastaje električna struja koja iritira susjedne dijelove membrane. Međutim, samo dio membrane u području sljedećeg Ranvierovog čvora može ući u stanje ekscitacije. Dakle, ekscitacija se širi preko membrane na skokovit (salativan) način od jednog presretanja Ranviera do drugog.
FUNKCIJE NEURONA
Materijal za izgradnju centralnog nervnog sistema i njegovih provodnika je nervno tkivo koje se sastoji od dve komponente - nervnih ćelija (neurona) i neuroglije. Glavni funkcionalni elementi CNS-a su neuroni: u tijelu životinja ih ima oko 50 milijardi, od kojih se samo mali dio nalazi u perifernim dijelovima tijela.
Neuroni čine 10-15% ukupnog broja ćelijskih elemenata u nervnom sistemu. Njegov glavni dio zauzimaju neuroglijalne ćelije. Kod viših životinja, u procesu postnatalne ontogeneze, diferencirani neuroni se ne dijele. (kao što se ranije mislilo). Neuroni se značajno razlikuju po obliku (piramidalni, okrugli, zvezdasti, ovalni), veličini (od 5 do 150 mikrona), broju procesa, ali imaju i zajednička svojstva.
U nekim dijelovima mozga postoje neuroni koji proizvode granule sekrecije mukoproteinske ili glikoproteinske prirode. Imaju i fiziološke karakteristike neurona i žljezdanih stanica. Ove ćelije se nazivaju neurosekretorne. Funkcija neurona je da percipiraju signale od receptora ili drugih nervnih ćelija, pohranjuju i obrađuju informacije i prenose nervne impulse drugim ćelijama - nervnim, mišićnim ili sekretornim. Shodno tome, postoji specijalizacija neurona. Podijeljeni su u 3 grupe: osjetljivi (senzorni, aferentni) neuroni, asocijativni (srednji, interkalarni) neuroni, motorni (efektorni) neuroni.
Neuroglia. Najveći dio nervnog tkiva čine glijalni elementi koji obavljaju pomoćne funkcije i ispunjavaju gotovo cijeli prostor između neurona. Anatomski, među njima se razlikuju neuroglijalne ćelije u mozgu (oligodendrociti i astrociti) i Schwannove ćelije u perifernom nervnom sistemu. Oligodendrociti i Schwannove ćelije formiraju mijelinske ovojnice oko aksona. Između glijalnih ćelija i neurona postoje praznine širine 15 - 20 nm, koje međusobno komuniciraju, formirajući intersticijski prostor ispunjen tekućinom. Kroz ovaj prostor dolazi do razmjene supstanci između neurona i glijalnih stanica, kao i opskrbe neurona kisikom i hranjivim tvarima difuzijom. Glijalne ćelije, po svemu sudeći, obavljaju samo potporne i zaštitne funkcije u CNS-u, a nisu, kako se očekivalo, njihov izvor ishrane ili čuvari informacija.
Glijalne ćelije se razlikuju od neurona po svojstvima membrane: pasivno reaguju na električnu struju, njihove membrane ne generišu propagirajući impuls. Postoje uske spojeve (područja niskog otpora) između neuroglijalnih ćelija koje pružaju direktnu električnu vezu. Membranski potencijal glijalnih ćelija je veći od potencijala neurona i uglavnom zavisi od koncentracije K+ jona u medijumu.
Kada se koncentracija K+ poveća tokom aktivne aktivnosti neurona u ekstracelularnom prostoru, jedan dio apsorbuju depolarizirani glijalni elementi. Ova puferska funkcija glije osigurava relativno konstantnu ekstracelularnu koncentraciju K+. Glia ćelije - astrociti - nalaze se između tijela neurona i zida kapilara, njihovi procesi su u kontaktu sa zidom potonjeg. Ovi perivaskularni procesi su elementi krvno-moždane barijere. Mikroglijalne stanice obavljaju fagocitnu funkciju, njihov broj naglo raste kada je oštećeno moždano tkivo.
Fiziologija sinapsi.
Hemijske sinapse imaju relativno širok sinaptički rascjep (20 - 50 nm) i visoku otpornost sinaptičkih membrana. Presinaptički nervni terminal sadrži veliki broj vezikula - sinaptičkih vezikula - oko 50 nm u prečniku, ispunjen medijatorom.
Mehanizam prijenosa ekscitacije u kemijskim ekscitatornim sinapsama. U sinapsama sa hemijskim prenosom, ekscitacija se prenosi pomoću posrednici(posrednici). Mediat ory - to su hemikalije koje obezbeđuju prenos ekscitacije u sinapsama. Medijatori se, ovisno o svojoj prirodi, dijele u nekoliko grupa:
monoamini(acetilholin, dopamin, norepinefrin, serotonin, itd.);
Amino kiseline(gama-aminobutirna kiselina - GABA, glutaminska kiselina, glicin, itd.);
neuropeptidi(supstanca P, endorfini, neurotenzin, ACTH, angiotenzin, vazopresin, somatostatin itd.). Posrednik u molekularnom obliku nalazi se u vezikulama presinaptičkog zadebljanja (sinaptički plak), gdje ulazi:
Iz perinuklearne regije neurona pomoću brzog aksonskog transporta (aksostruja);
Zbog sinteze medijatora koji se javlja u sinaptičkim terminalima iz proizvoda njegovog cijepanja;
Zbog ponovnog hvatanja neurotransmitera iz sinaptičke pukotine u nepromijenjenom obliku.
Raznovrsnost medijatorskih sistema. Dale princip: jedan neuron, po pravilu, sintetiše i koristi jedan posrednik u svim svojim terminalima. Moguće je da neuron koristi nekoliko medijatora (komičari) , ali očigledno u istoj kombinaciji (ovaj princip je trenutno sporan, pogledajte dolje za sisteme medijatora) . Kao posljedica Daleovog principa, mogu se smatrati sljedeće odredbe:
1) Znak sinaptičkog djelovanja ne određuje medijator, već svojstva receptora na postsinaptičkoj ćeliji.
2) Receptori na stanicama koji su postsinaptični za jedan presinaptički neuron mogu se farmakološki razlikovati i mogu kontrolirati različite ionske kanale.
3) Jedna postsinaptička ćelija može imati više od jednog tipa receptora za dati medijator, a svaki od ovih receptora može kontrolirati drugačiji mehanizam jonske provodljivosti.
Zbog ova tri svojstva, ćelije mogu vršiti suprotna sinaptička dejstva kako na različite postsinaptičke ćelije tako i na istu.
Do sada identifikovani medijatori kod životinja i ljudi čine prilično heterogenu grupu supstanci. Monoamini: acetilholin, dopamin, norepinefrin, serotonin (5-hidroksitriptamin, 5-HT), histamin. Aminokiseline: gama-aminobuterna kiselina (GABA), glutaminska kiselina, glicin, taurin, itd. Neuropeptidi: metenkefalin, leuenkefalin, endorfin, oksitocin ,vazopresin i sl . Odgovarajuće sinapse nazivaju se npr. holinergički, serotonergički, noradrenergic itd. Veliki broj medijatora odgovara velikom broju postsinaptičkih receptora: holinergičkih receptora , adrenoreceptori, GABA receptori itd. Unutar svake grupe receptora postoji niz njihovih podtipova, na primjer, nikotinski i muskarinski acetilkolinski receptori (H-XR i M-XR, respektivno).
Podjela receptora se vrši na osnovu razlika u njihovim farmakološkim svojstvima: različiti agonisti (supstance koje oponašaju dejstvo posrednika) i antagonisti (supstance koje sprečavaju ispoljavanje dejstva medijatora). Na primjer, za N-ChR, agonist je nikotin , antagonisti - tubokurarin (izolovan iz otrova kurare), bungarotoksin (izolovan iz otrova zmije iz roda Bungarus). Postoje tri tipa glutamatnih receptora zasnovanih na agonistima: quisqualate (AMPA-tip), kainate I NMDA (N-metil-D-aspartat) -tip. Neki posrednički receptori (posebno, adrenoreceptori i receptori za mnoge neuropeptide) nisu povezani sa ionskim kanalima (jonotropni receptori) , ali sa membranskim enzimom (metabotropni receptori) , na primjer, adenilat ciklaza . Potonji, jednom aktiviran od strane posrednika, katalizira transformaciju mnogih molekula adenozin trifosfat (ATP) ciklično adenozin monofosfat (cAMP) - mehanizam za pojačanje. CAMP, kao drugi glasnik, posebno aktivira mnoge enzime u ćeliji protein kinaze i na taj način stimuliše ćelijski metabolizam. cAMP je uništen fosfodiesteraza. Sistem adenilat ciklaze je sličan sistemu gvanilat ciklaze, sistemu fosfolipaze C, itd. (Sl. 14). Formiranje cGMP iz necikličkog oblika katalizira gvanilat ciklaza čiju aktivnost stimulira dušikov oksid. Molekuli potonjeg nastaju tokom deaminacije arginina i stvaranja citrulina pod djelovanjem sintaza dušikovog oksida. Aktivnost ovog enzima je zauzvrat regulirana kompleksom Ca 2+ -kalmodulin. Tako, na primjer, glutamatni receptori, koji pokreću dolazni protok Ca 2+ jona, kontroliraju fluktuacije koncentracije cGMP u citoplazmi neurona. U mozgu kralježnjaka, aktivnost sintaze dušikovog oksida nalazi se u malom mozgu, kvadrigemini, striatumu i olfaktornoj lukovici.
Mehanizam hemijske sinapse: prilikom depolarizacije presinaptičkog terminala (prouzrokovane PD ili veštački), iz okoline u njega ulaze ioni Ca 2+ koji stimulišu proces egzocitoza - pražnjenje vezikula u sinaptičku pukotinu.
Prikazani su karakteristični sinaptički i vezikularni proteini sa njihovim navodnim receptorima i funkcijama. Odvojeni dijelovi vezikularne membrane postulirani su za učvršćivanje vezikula na citoskelet), pričvršćivanje vezikularne membrane na presinaptičku membranu i oslobađanje medijatora kroz nastalu poru. Molekularni mehanizmi vezivanja vezikula za presinaptičku membranu i formiranje pora su vjerovatno različiti. Neki od proteina su mete neurotoksina koji mijenjaju oslobađanje neurotransmitera. Na primjer, struktura vezikularnih sinaptobrevin proteina (VAMPs) je poremećena tetanusom i botulinum toksinima; latrotoksin paukovog otrova se veže za proteine presinaptičke membrane neureksine i pojačava pražnjenje vezikula.
1. Sinapsini su proteini povezani s vezikulama koji vjerojatno povezuju sinaptičke vezikule sa citoskeletom nervnog završetka. 2. Pričvršćivanje, formiranje pora i pražnjenje vezikula posredovani su interakcijama (označeno strelicama) različitih vezikularnih i membranskih proteina. Na primjer, vezikularni proteini (sinaptotagmin i sinaptobrevini) i proteini plazma membrane nervnog završetka (sintaksini i neureksini) su uključeni u formiranje kompleksa vezivanja. 3. Još uvijek nije jasno koji proteini - plazmatski ili vezikularni formiraju fuzijsku poru. Vjerovatno su to sinaptofizin (koji ima druge funkcije) i protein plazma membrane fizofilin. 4. Rab proteini mogu biti uključeni u transport vezikula u ćeliji i u njihovom vezivanju za membranu živčane stanice. Vezikularni transporteri igraju ulogu u akumulaciji neurotransmitera u sinaptičkim vezikulama.
Istovremeno, Ca 2+ počinje da se uklanja iz citoplazme na nekoliko načina: vezivanje proteina, mitohondrijski unos i aktivni transport. Izlaz medijatora zavisi od depolarizacije terminala i iznosi oko 100 - 200 vezikula, od kojih svaka sadrži jedan deo (kvantni), što odgovara približno 10 4 molekula. Molekuli medijatora difundiraju do postsinaptičke membrane, gdje stupaju u interakciju sa postsinaptičkim membranskim receptorima koji regulišu stanje jonskih kanala. Ova regulacija može biti direktna (kao, na primjer, u neuromuskularnom spoju skeletnih mišića kralježnjaka) ili uključivati aktivaciju sistema sekundarno intracelularni medijatori (G proteini, cAMP).
U prvom slučaju se prenose brzi startni signali, u drugom se provode sporiji dugoročni efekti. Smjer promjene potencijala postsinaptičke membrane (depolarizacija ili hiperpolarizacija) ovisi uglavnom o tome čijim otvaranjem kanala upravljaju postsinaptički receptori. Neki od molekula medijatora mogu stupiti u interakciju presinaptički receptori, što dovodi do promjene MP nervnog terminala i, shodno tome, količine oslobođenog medijatora (povratna informacija). Sinaptički rascjep se čisti od medijatora na različite načine: deaktivacijom, hidrolizom, ponovnim unosom u presinaptički završetak, difuzijom, hvatanjem glijalnih stanica. Glavni dio sinaptičko kašnjenje - vrijeme od dolaska nervnog impulsa do razvoja postsinaptičkog odgovora (0,2-0,5 ms) pada na proces lučenja neurotransmitera. Hemijska sinapsa omogućava prijenos signala samo od presinaptičkog neurona do postsinaptičkog.
Uz čestu ritmičku stimulaciju u hemijskim sinapsama, povećava se (olakšanje) a zatim i slabljenje (depresija) prenos, tj. raste, a zatim pada amplituda postsinaptičkih potencijala. Ove pojave su uglavnom određene promjenama presinaptičke veze. Oni imaju poseban razvoj u nekim sinapsama CNS-a, gdje djeluju kao faktori sinaptičke plastičnosti. Prisustvo različitih tipova receptora u postsinaptičkoj membrani može uzrokovati razvoj ovih pojava odvojeno, kao što su dugotrajna potenciranje i dugotrajna depresija.
Hemijske inhibitorne sinapse. Ove sinapse, u smislu mehanizma prijenosa ekscitacije, slične su sinapsama ekscitatornog djelovanja. U inhibitornim sinapsama medijator (na primjer, glicin) stupa u interakciju s receptorima na subsinaptičkoj membrani i otvara kloridne kanale u njoj, što dovodi do kretanja kloridnih jona duž gradijenta koncentracije u ćeliju i razvoja hiperpolarizacije na podsinaptičkoj membrani. . Postoji tzv inhibicijski postsinaptički potencijal(TPSP).
Ranije se vjerovalo da svaki medijator odgovara specifičnoj reakciji postsinaptičke stanice - ekscitaciji ili inhibiciji u ovom ili onom obliku. Sada je utvrđeno da jedan medijator najčešće odgovara ne jednom, već nekoliko različitih receptora. Na primjer, acetilholin u neuromuskularnim sinapsama skeletnih mišića djeluje na H-holinergičke receptore (osjetljive na nikotin), koji otvaraju široke kanale za natrijum (i kalij), koji stvara EPSP (PKP) U vago-srčanim sinapsama isti acetilholin djeluje na M-holinergički receptori (osetljivi na muskarin), koji otvaraju selektivne kanale za jone kalijuma, pa se ovde generiše inhibitorni postsinaptički potencijal (IPSP). Shodno tome, ekscitatorna ili inhibitorna priroda djelovanja medijatora određena je svojstvima subsinaptičke membrane (tačnije, vrstom receptora), a ne samim medijatorom.
Fiziološka svojstva hemijskih sinapsi. Sinapse sa hemijskim prenosom ekscitacije imaju niz zajedničkih svojstava:
Ekscitacija kroz sinapse se provodi samo u jednom smjeru (jednostrano). To je zbog strukture sinapse: medijator se oslobađa samo iz presinaptičkog zadebljanja i stupa u interakciju s receptorima podsinaptičke membrane;
Prenos ekscitacije kroz sinapse je sporiji nego kroz nervno vlakno – sinaptičko kašnjenje;
Prijenos ekscitacije vrši se uz pomoć posebnih kemijskih medijatora - medijatora;
U sinapsama se transformiše ritam ekscitacije;
Sinapse imaju nisku labilnost;
Sinapse su veoma umorne;
Sinapse su veoma osetljive na hemijske (uključujući farmakološke) supstance.
Ekscitatorne električne sinapse. Pored sinapsi sa hemijskim prenosom ekscitacije, sinapse sa električnim prenosom nalaze se pretežno u centralnom nervnom sistemu (CNS). Ekscitatorne električne sinapse karakterizira vrlo uska sinaptička pukotina i vrlo niska otpornost susjednih pre- i postsinaptičkih membrana, što osigurava efikasan prolaz lokalnih električnih struja. Nizak otpor, u pravilu, povezan je s prisustvom poprečnih kanala koji prelaze obje membrane, odnosno idu od ćelije do ćelije (kontakt praznina). Kanale formiraju proteinski molekuli (polumolekule) svake od kontaktnih membrana, koje su komplementarno povezane. Ova struktura je lako prohodna za električnu struju.
Dijagram prijenosa ekscitacije u električnoj sinapsi: struja uzrokovana presinaptičkim akcijskim potencijalom iritira postsinaptičku membranu, gdje se javljaju EPSP i akcijski potencijal. Poprečni kanali ujedinjuju ćelije ne samo električni, već i hemijski, jer su prohodni za mnoga jedinjenja male molekularne težine. Stoga se ekscitatorne električne sinapse s poprečnim kanalima formiraju, u pravilu, između stanica istog tipa (na primjer, između stanica srčanog mišića).
Opšta svojstva ekscitatornih električnih sinapsi su:
Brzina (značajno premašuje onu u hemijskim sinapsama);
Slabost efekata tragova tokom prenosa ekscitacije (kao rezultat toga, sumiranje uzastopnih signala u njima je praktično nemoguće);
Visoka pouzdanost prijenosa pobude.
Ekscitatorne električne sinapse mogu nastati pod povoljnim uslovima i nestati pod nepovoljnim. Na primjer, ako je jedna od ćelija u kontaktu oštećena, njene električne sinapse s drugim stanicama su eliminirane. Ovo svojstvo se naziva plastičnost. Električne sinapse mogu biti jednostrane ili bilateralne.
Električna inhibitorna sinapsa. Uz električne sinapse ekscitatornog djelovanja, mogu se pojaviti i električne inhibicijske sinapse. Inhibicijski učinak nastaje djelovanjem struje uzrokovane akcionim potencijalom presinaptičke membrane. Presinaptički potencijal uzrokuje značajnu hiperpolarizaciju segmenta, a hiperpolarizirajuća struja trenutno inhibira stvaranje akcionog potencijala u početnom segmentu aksona.
IN mješovite sinapse presinaptički akcijski potencijal stvara struju koja depolarizira postsinaptičku membranu tipične kemijske sinapse, gdje se pre- i postsinaptička membrana ne prianjaju čvrsto jedna za drugu. Stoga, u ovim sinapsama, hemijski prijenos služi kao neophodan mehanizam za jačanje.
Trofički uticaji koji se prenose kroz sinapse. Osim što prenose ekscitatorne i inhibitorne signale koji su funkcionalno važni, sinapse pružaju trophic (tj. utiču na rast i diferencijaciju) interakcije kontaktnih ćelija, ostvarene uz pomoć trofičkih faktora proteinske prirode, verovatno akumulirane i u vezikulama. Ovi faktori obezbeđuju metaboličko održavanje potrebne strukture i svojstava ovih ćelija. Bilateralne trofičke interakcije su predložene u svim sinapsama, ali su proučavane uglavnom u neuromuskularnim sinapsama skeleta kralježnjaka. Denervacija mišića dovodi do gubitka mišićnih vlakana diferencijacije postignute u ontogenezi.
postsinaptički procesi. Promjene u potencijalu postsinaptičke membrane kao rezultat aktivacije sinapse nazivaju se sinaptički potencijal. Ekscitatorni postsinaptički potencijal (VPSP) javlja se u depolarizirajućim sinapsama i nastaje zbog istovremenog povećanja permeabilnosti membrane za Na+ i K+ ione. Struje koje nastaju u ovom slučaju su suprotno usmjerene (natrij - unutar ćelije, kalij - van). MF se pomiče prema depolarizaciji do vrijednosti jednake polovini sume ravnotežnih potencijala E Na i E K . Inhibicijski postsinaptički potencijali (IPSP) su hiperpolarizacijske promjene u magnetskom polju (do -80-90 mV) i nastaju zbog otvaranja kanala za K+ ione (koji napuštaju ćeliju), ili Cl - ione (ulaze u ćeliju), ili za oba jona na isto vrijeme. Amplituda sinaptičkih potencijala zavisi od količine oslobođenog medijatora (broja kvanta) i, stoga, ove reakcije su postepeno za razliku od PD. Ovo amplitudno kodiranje frekvencijskog signala se provodi u postsinaptičkom neuronu, sa izuzetkom njegove aksonalne regije, u kojoj dolazi do povratka na frekvencijsko kodiranje zbog AP koji se širi duž aksonskog vlakna.
Depolarizacija koju stvara EPSP može značajno premašiti, dostići ili ostati ispod praga ekscitacije (označeno tačkama). Amplituda hiperpolarizacije usled razvoja IPSP zavisi od frekvencije presinaptičke AP i utiče na učestalost pozadinske aktivnosti postsinaptičke ćelije.
Jer smjer jonskih struja ovisi o gradijentu elektrohemijskog potencijala datog jona, tada se amplituda i polaritet sinaptičkog signala mijenjaju s promjenom magnetnog polja. Naziva se njegova vrijednost na kojoj se mijenja znak sinaptičkog djelovanja potencijal preokreta. Širenje sinaptičkih potencijala je samo zbog fizičkih svojstava stanične membrane i stoga se javlja sa slabljenjem (smanjenjem amplitude). Sinaptički potencijali koji nastaju u različitim sinapsama mogu međusobno komunicirati, poštujući pravila algebarskog zbrajanja. Depolarizacija uzrokovana sumiranjem EPSP dovodi MP bliže graničnoj vrijednosti za pobuđivanje. Naprotiv, nametanje TSSP-a ga udaljava. Stepen depolarizacije ili hiperpolarizacije membrane zavisi od „konfrontacije“ između jonskih provodljivosti i struja koje se aktiviraju tokom EPSP i IPSP. Glavnu ulogu u integraciji signala koji dolaze u ćeliju imaju posebni dijelovi neuronske membrane: čvorovi grananja dendritnog stabla, somatska membrana i brežuljak aksona. Ekscitatorne sinapse su najčešće lokalizovane na dendritskoj membrani, dok se inhibitorne sinapse nalaze u pravilu na somatskoj membrani. Konačna integracija svih ulaza događa se na kolikulusu aksona. Budući da se postsinaptički procesi odvijaju na različitim udaljenostima od zone okidača i propagiraju pasivno, njihov doprinos integrativnom izlazu ovisit će o lokalizaciji sinaptičkih kontakata.
Na ovu interakciju utiču geometrijski odnosi između ekscitatornih i inhibitornih sinapsi lociranih u različitim delovima dendrita, kao i karakteristike propagacije elektrotonične struje duž ovih dendrita. U ovom slučaju, u postsinaptičkom neuronu, tokom generisanja AP, mogu se javiti efekti sabiranja i oduzimanja ulaznih signala koji imaju iste ili različite (ekscitatorne i inhibitorne) modalitete.
Prijenos signala nepulsirajućim neuronima. Neke nervne ćelije ne stvaraju AP u fiziološkim uslovima (ova sposobnost, tipična za neurone, manifestuje se nakon određenih vrsta hemijskog izlaganja). Primjeri takvih ćelija u kralježnjaka su fotoreceptori i granularne ćelije olfaktornih lukovica. Međutim, ovi neuroni zadržavaju obrazac: neurotransmiter se oslobađa tek nakon depolarizacije presinaptičke ćelije.
Ekscitacija, koja je nastala u jednom dijelu membrane ekscitabilne stanice, ima sposobnost širenja. Dugi proces neurona - aksona (nervno vlakno) obavlja u tijelu specifičnu funkciju provođenja ekscitacije na velike udaljenosti.
Zakoni ekscitacijeduž nervnih vlakana
Zakon anatomskog i fiziološkog kontinuiteta - ekscitacija se može širiti duž nervnog vlakna samo ako je morfološki i funkcionalno netaknuta.
Zakon bilateralnog provođenja ekscitacije- ekscitacija koja se javlja u jednom dijelu živca širi se u oba smjera od mjesta nastanka. U tijelu, ekscitacija se uvijek širi duž aksona od tijela ćelije (ortodromski).
Zakon izolovanog ponašanja- ekscitacija koja se širi duž vlakna koje je dio živca ne prenosi se na susjedna nervna vlakna.
Obrasci lokalnogi širenje uzbuđenja
Elektrotonični potencijal (lokalna ekscitacija)
• širi se duž nervnih vlakana sa slabljenjem (sa dekrement), tj. amplituda lokalnog odgovora brzo opada sa povećanjem udaljenosti od mjesta njegovog pojavljivanja;
• zbog slabljenja, lokalni odziv se proteže na kratke udaljenosti (ne više od 2 cm);
• lokalna ekscitacija se širi pasivno, bez trošenja ćelijske energije;
• mehanizam širenja lokalne pobude sličan je širenju električne struje u provodnicima; Ova vrsta razmnožavanja se zove elektrotonski.
Akcioni potencijal (širenje ekscitacije)
• širi se duž nervnih vlakana bez slabljenja, amplituda akcionog potencijala je ista na bilo kojoj udaljenosti od mesta njegovog nastanka;
• udaljenost do koje se proteže akcioni potencijal ograničena je samo dužinom nervnog vlakna;
• propagacija akcionog potencijala je aktivan proces tokom kojeg se menja stanje jonskih kanala vlakana, potrebna je ATP energija da bi se obnovili transmembranski jonski gradijenti;
• mehanizam provođenja akcionog potencijala je složeniji od mehanizma propagacije lokalne ekscitacije.
Mijelinska i nemijelinizirana nervna vlakna
mijelinska vlakna. Neka nervna vlakna prolaze kroz mijelinizaciju tokom embriogeneze: lemociti (Schwannove ćelije) prvo dodiruju akson, a zatim ga obavijaju (sl. 1, A, B). Membrana lemocita obavija se oko aksona poput rolne, formirajući višeslojnu spiralu (mijelinski omotač) (Sl. 1, C, D). Mijelinska ovojnica nije kontinuirana - duž cijele dužine nervnog vlakna na jednakoj udaljenosti jedan od drugog, u njemu su mali prekidi (presreće Ranvier). U području presjeka, akson je lišen mijelinske ovojnice.
nemijelinizirana vlakna. Mijelinizacija ostalih vlakana završava se u ranim fazama embrionalnog razvoja. Jedan ili više aksona je uronjeno u lemocit; potpuno ili djelimično ih okružuje, ali ne formira višeslojni mijelinski omotač (slika 1e).
Mehanizam provođenja ekscitacije duž nemijeliniziranih nervnih vlakana
U mirovanju, cijela unutrašnja površina membrane nervnog vlakna nosi negativan naboj, a vanjska strana membrane je pozitivna. Električna struja između unutrašnje i vanjske strane membrane ne teče, jer lipidna membrana ima visok električni otpor.
Tokom razvoja akcionog potencijala dolazi do reverzije naelektrisanja u pobuđenom području membrane (slika 2, A). Električna struja počinje da teče na granici pobuđenog i nepobuđenog preseka (slika 2, B). Električna struja iritira najbliži dio membrane i dovodi ga u stanje ekscitacije (slika 2, C), dok se prethodno pobuđeni dijelovi vraćaju u stanje mirovanja (slika 2, D). Tako talas ekscitacije pokriva sve nove delove membrane nervnih vlakana.
Mehanizam provođenja ekscitacije duž mijelinskih nervnih vlakana
U mijeliniziranom nervnom vlaknu, dijelovi membrane prekriveni mijelinskom ovojnicom nisu ekscitabilni; ekscitacija se može dogoditi samo u područjima membrane koja se nalaze u području Ranvierovih presjeka.Sa razvojem AP dolazi do preokreta membranskog naboja u jednom od Ranvierovih čvorova (slika 3, A). Između elektronegativnog i elektropozitivnog dijela membrane nastaje električna struja koja iritira susjedne dijelove membrane (slika 3, B). Međutim, samo dio membrane u području sljedećeg Ranvierovog čvora može ući u stanje ekscitacije (slika 3c). Dakle, ekscitacija se širi preko membrane na skokovit (salativan) način od jednog presretanja Ranviera do drugog.
Klasifikacija nervnih vlakana
Nervna vlakna variraju u prečniku i stepenu mijelinizacije. Što je veći prečnik nervnog vlakna i stepen mijelinizacije, to je veća brzina provođenja ekscitacije. Vlakna s različitim brzinama provodljivosti obavljaju različite fiziološke funkcije. Nervna vlakna se dijele na 6 tipova, čije su karakteristike date u tabeli. 4.1.
Tabela 4.1. Vrste nervnih vlakana, njihova svojstva i funkcionalna namjena
Prečnik (µm) |
mijelinizacija |
Brzina provodljivosti (m/s) |
Funkcionalna namjena |
|
Motorna vlakna somatskog NS; senzorna vlakna proprioreceptora |
||||
Senzorna vlakna kožnih receptora |
||||
Senzorna vlakna proprioreceptora |
||||
Senzorna vlakna termoreceptora, nociceptora |
||||
Preganglijska vlakna simpatičkog NS-a |
||||
nedostaje |
Postganglijska vlakna simpatičkog NS; senzorna vlakna termoreceptora, nociceptora, nekih mehanoreceptora |
Nervna vlakna svih grupa imaju zajednička svojstva:
• nervna vlakna su praktično neumorna;
• nervna vlakna imaju visoku labilnost, odnosno mogu reproducirati akcioni potencijal sa vrlo visokom frekvencijom.

