20.01.2022
Μη μυελινωμένες και μυελινωμένες νευρικές ίνες. Η έννοια της "νευρικής ίνας"
Μια νευρική ίνα είναι μια επιμήκης διαδικασία νευρώνων που καλύπτονται με λεμοκύτταρα και ένα περίβλημα μυελίνης ή μη. Η κύρια λειτουργία του είναι η αγωγή των νευρικών ερεθισμάτων. Στο περιφερικό και κεντρικό νευρικό σύστημα, κυριαρχούν οι πολφώδεις (μυελινωμένες) νευρικές ίνες που νευρώνουν τους σκελετικούς μύες. Οι ίνες που δεν έχουν θήκη ονομάζονται γυμνοί αξονικοί κύλινδροι.
Η νευρική ίνα βασίζεται στη διαδικασία του νευρώνα, ο οποίος σχηματίζει ένα είδος άξονα. Εξωτερικά, περιβάλλεται από ένα περίβλημα μυελίνης με μια βιομοριακή λιπιδική βάση, που αποτελείται από μεγάλο αριθμό στροφών μεσαξόνης, το οποίο τυλίγεται σπειροειδώς γύρω από τον νευρωνικό άξονα. Έτσι, λαμβάνει χώρα μυελίνωση των νευρικών ινών.
Οι μυελινωμένες νευρικές ίνες του περιφερικού συστήματος καλύπτονται επιπλέον από πάνω από βοηθητικά κύτταρα Schwann που υποστηρίζουν τον άξονα και τροφοδοτούν το σώμα του νευρώνα. Η επιφάνεια της πολτοποιημένης μεμβράνης έχει διαστήματα - παρεμβολές του Ranvier, σε αυτά τα σημεία ο αξονικός κύλινδρος συνδέεται με την εξωτερική μεμβράνη Schwann.
Το στρώμα μυελίνης δεν έχει ηλεκτρικά αγώγιμες ιδιότητες, έχουν τομές. Η διέγερση εμφανίζεται στο διάστημα Ranvier που βρίσκεται πιο κοντά στη θέση έκθεσης σε ένα εξωτερικό ερέθισμα. Η ώθηση μεταδίδεται απότομα, από τη μια αναχαίτιση στην άλλη, αυτό παρέχει υψηλή ταχύτητα διάδοσης της ώθησης.
Οι νευρικές ίνες της μυελίνης ρυθμίζουν το μεταβολισμό στον μυϊκό ιστό, έχουν υψηλή αντίσταση στο βιοηλεκτρικό ρεύμα.
Τα κενά του Ranvier δημιουργούν και ενισχύουν παρορμήσεις. Οι ίνες του κεντρικού νευρικού συστήματος δεν έχουν μεμβράνη Schwann· αυτή η λειτουργία εκτελείται από την ολιγοδενδρογλοία.
Οι αμυελιωμένοι ιστοί έχουν αρκετούς αξονικούς κυλίνδρους, δεν έχουν στρώμα μυελίνης και παρεμβολές, καλύπτονται με κύτταρα Schwann από πάνω, σχηματίζονται χώροι σαν σχισμή μεταξύ τους και των κυλίνδρων. Οι ίνες έχουν κακή μόνωση, επιτρέπουν τη διάδοση μιας ώθησης από τη μια διεργασία ενός νευρώνα στην άλλη, επαφή με το περιβάλλον καθ' όλη τη διάρκεια, η ταχύτητα αγωγής των παλμών είναι πολύ χαμηλότερη από αυτή των πολτοειδών ινών, ενώ το σώμα χρειάζεται περισσότερη ενέργεια.
Από τις πολφώδεις και μη σαρκώδεις διεργασίες των νευρώνων σχηματίζονται μεγάλοι νευρικοί κορμοί, οι οποίοι με τη σειρά τους διακλαδίζονται σε μικρότερες δέσμες και καταλήγουν σε νευρικές απολήξεις (υποδοχέας, κινητήρας, συνάψεις).
Οι νευρικές απολήξεις είναι το άκρο των μυελινωμένων και μη μυελινωμένων νευρικών ινών, οι οποίες σχηματίζουν ενδονευρικές επαφές, υποδοχείς και κινητικές απολήξεις.
Αρχές ταξινόμησης

Διαφορετικοί τύποι νευρικών ινών έχουν άνισο ρυθμό αγωγής διεγερτικών ερεθισμάτων, αυτό εξαρτάται από τη διάμετρό τους, τη διάρκεια του δυναμικού δράσης και τον βαθμό μυελίνωσης. Υπάρχει μια ευθέως ανάλογη σχέση μεταξύ της ταχύτητας και της διαμέτρου της ίνας.
Δομική-λειτουργική μέθοδος ταξινόμησης των νευρικών ινών Erlanger-Gasser ανάλογα με την ταχύτητα αγωγής των νευρικών παλμών:
- Ομάδα Α μυελινωμένη νευρική ίνα: α, β, Υ και δ. Η μεγαλύτερη διάμετρος και το παχύτερο κέλυφος είναι ιστοί α - 20 μικρά, έχουν καλή ταχύτητα αγωγής παλμού - 120 m / s. Αυτοί οι ιστοί νευρώνουν την πηγή διέγερσης από τη στήλη του νωτιαίου μυελού στους υποδοχείς των σκελετικών μυών, τους τένοντες και είναι υπεύθυνοι για τις απτικές αισθήσεις.
Οι υπόλοιποι τύποι ινών έχουν μικρότερη διάμετρο (12 μικρά), την ταχύτητα της ώθησης. Αυτοί οι ιστοί μεταδίδουν σήματα από εσωτερικά όργανα, πηγές πόνου στο ΚΝΣ.
- Οι ίνες μυελίνης της ομάδας Β ανήκουν. Η συνολική ταχύτητα αγωγής παλμών είναι 14 m/s, το δυναμικό δράσης είναι 2 φορές μεγαλύτερο από αυτό των ινών της ομάδας Α. Το περίβλημα μυελίνης εκφράζεται ελάχιστα.
- Οι μη μυελινωμένες ίνες της ομάδας C έχουν πολύ μικρή διάμετρο (0,5 microns) και ταχύτητα διέγερσης (6 m / s). Αυτοί οι ιστοί νευρώνουν το συμπαθητικό νευρικό σύστημα. Αυτή η ομάδα περιλαμβάνει επίσης ίνες που διεξάγουν ώσεις από τα κέντρα του πόνου, του κρύου, της ζέστης και της πίεσης.
Οι διεργασίες των νευρώνων χωρίζονται σε προσαγωγούς και απαγωγούς. Ο πρώτος τύπος εξασφαλίζει τη μετάδοση των παλμών από τους υποδοχείς των ιστών στο κεντρικό νευρικό σύστημα. Ο δεύτερος τύπος μεταδίδει τη διέγερση από το κεντρικό νευρικό σύστημα στους υποδοχείς των ιστών.
Λειτουργική ταξινόμηση των νευρικών ινών του προσαγωγού τύπου κατά Lloyd-Hunt:
Απομυενιλοποίηση
Η διαδικασία απομυελίνωσης των νευρικών ινών είναι μια παθολογική βλάβη στο περίβλημα της μυελίνης, η οποία προκαλεί δυσλειτουργία των ιστών. Η παθολογία προκαλείται από φλεγμονώδεις διεργασίες, μεταβολικές διαταραχές, νευρολοίμωξη, δηλητηρίαση ή ισχαιμία των ιστών. Η μυελίνη αντικαθίσταται από ινώδεις πλάκες, με αποτέλεσμα την εξασθενημένη αγωγή των παλμών.
Ο πρώτος τύπος απομυελίνωσης είναι η μυελοπάθεια που προκαλείται από αυτοάνοσες αντιδράσεις του σώματος, η νόσος Canavan, το σύνδρομο Guillain-Barré, η αμυοτροφία Charcot-Marie-Tooth.
Ο δεύτερος τύπος είναι η μυελινοκλαστία. Η παθολογία χαρακτηρίζεται από κληρονομική προδιάθεση για την καταστροφή του ελύτρου της μυελίνης (νόσος Binswanger).
Απομυελινωτικές ασθένειες

Οι ασθένειες που οδηγούν στην καταστροφή του ελύτρου της μυελίνης είναι τις περισσότερες φορές αυτοάνοσης φύσης, μια άλλη αιτία μπορεί να είναι η θεραπεία με αντιψυχωσικά ή μια κληρονομική προδιάθεση. Η καταστροφή της λιπιδικής στιβάδας προκαλεί μείωση της ταχύτητας αγωγής των ερεθισμάτων.
Οι ασθένειες χωρίζονται σε αυτές που επηρεάζουν το κεντρικό νευρικό σύστημα και σε παθολογίες που βλάπτουν το περιφερειακό δίκτυο. Ασθένειες που επηρεάζουν τη λειτουργία του κεντρικού νευρικού συστήματος:
- Η μυελοπάθεια του νωτιαίου μυελού εμφανίζεται ως αποτέλεσμα της συμπίεσης των ινών της μυελίνης από μεσοσπονδύλιες κήλες, όγκους, θραύσματα οστών, μετά. Σε ασθενείς, η ευαισθησία και η μυϊκή δύναμη στην πληγείσα περιοχή μειώνονται, εμφανίζεται πάρεση των χεριών ή των ποδιών, διαταράσσεται η εργασία των εντέρων και του ουροποιητικού συστήματος και αναπτύσσεται ατροφία των μυών των κάτω άκρων.
- Η λευκοδυστροφία του εγκεφάλου προκαλεί βλάβη στη λευκή ουσία. Οι ασθενείς έχουν εξασθενημένο συντονισμό των κινήσεων, δεν μπορούν να κρατήσουν ισορροπία. Αναπτύσσεται μυϊκή αδυναμία, εμφανίζονται ακούσιοι σπασμοί, νευρικό τικ. Σταδιακή επιδείνωση της μνήμης, των πνευματικών ικανοτήτων, της όρασης και της ακοής. Στα τελευταία στάδια εμφανίζεται τύφλωση, κώφωση, πλήρης παράλυση και δυσκολία στην κατάποση τροφής.
- Η μικροεστιακή λευκοεγκεφαλοπάθεια του εγκεφάλου προσβάλλει συχνότερα άνδρες άνω των 60 ετών. Οι κύριοι λόγοι είναι η αρτηριακή υπέρταση και η κληρονομική προδιάθεση. Οι ασθενείς έχουν μειωμένη μνήμη και προσοχή, υπάρχει λήθαργος, δυσκολία στην ομιλία. Το βάδισμα επιβραδύνεται, ο συντονισμός των κινήσεων διαταράσσεται, εμφανίζεται ακράτεια ούρων, είναι δύσκολο για τον ασθενή να καταπιεί την τροφή.
- Το σύνδρομο ωσμωτικής απομυελίνωσης χαρακτηρίζεται από τη διάσπαση των περιβλημάτων μυελίνης στους εγκεφαλικούς ιστούς. Οι ασθενείς έχουν διαταραχή της ομιλίας, συνεχές αίσθημα υπνηλίας, κατάθλιψη ή ευερεθιστότητα, αλαλία, πάρεση όλων των άκρων. Στα αρχικά στάδια της νόσου, η διαδικασία απομυελίνωσης είναι αναστρέψιμη.
- Η σκλήρυνση κατά πλάκας εκδηλώνεται με μούδιασμα ενός ή δύο άκρων, μερική ή πλήρη απώλεια όρασης, πόνο κατά την κίνηση των ματιών, ζάλη, κόπωση, τρόμο των άκρων, διαταραχή του συντονισμού των κινήσεων, μυρμήγκιασμα σε διάφορα μέρη του σώματος.
- Η νόσος του Devic είναι μια φλεγμονώδης αυτοάνοση νόσος που επηρεάζει το οπτικό νεύρο και το νωτιαίο μυελό. Τα συμπτώματα περιλαμβάνουν ποικίλου βαθμού διαταραχή της όρασης, μέχρι τύφλωση, παραπάρεση, τετραπάρεση, διαταραχή της λειτουργίας των πυελικών οργάνων.
Τα συμπτώματα των ασθενειών εξαρτώνται από την περιοχή της βλάβης στις ίνες της μυελίνης. Μπορείτε να αναγνωρίσετε τη διαδικασία απομυελίνωσης χρησιμοποιώντας υπολογιστική τομογραφία, θεραπεία μαγνητικού συντονισμού. Ενδείξεις βλάβης στο περιφερικό νευρικό σύστημα εντοπίζονται στην ηλεκτρομυογραφία.
Μυελίνωση των προσαγωγών οπτικών οδώναρχίζει στο πλάγιο γεννητικό σώμα τον πέμπτο μήνα της κύησης και τελειώνει με τη γέννηση στην ακανθώδη πλάκα. Τα ολιγοδενδροκύτταρα που είναι υπεύθυνα για τη μυελίνωση των ινών του ΚΝΣ συνήθως απουσιάζουν στον αμφιβληστροειδή. Ιστολογικές μελέτες επιβεβαίωσαν την παρουσία υποτιθέμενων ολιγοδενδροκυττάρων και μυελίνης στις περιοχές των μυελινωμένων νευρικών ινών και την απουσία τους εκτός αυτών των περιοχών.
Στο αυτοψία ινών μυελίνηςο αμφιβληστροειδής ανιχνεύεται στο 1% περίπου των οφθαλμών και στο 0,3-0,6% των οφθαλμικών ασθενών κατά τη διάρκεια μιας εξέτασης ρουτίνας.
μυελινωμένες νευρικές ίνεςεμφανίζονται συνήθως ως διαμήκη ραβδωτές δεσμίδες στους άνω και κάτω πόλους του δίσκου. Ταυτόχρονα, λόγω της προεξοχής των αλλαγμένων τμημάτων του δίσκου και της επικάλυψης των άκρων του δίσκου και των υποκείμενων αιμοφόρων αγγείων από ίνες, η ανωμαλία μπορεί να προσομοιώσει το οίδημα του οπτικού δίσκου.
Απομακρυσμένα σχηματίζονται οι ίνες ακανόνιστο σχήμα ανεμιστήρα. Εντός της ζώνης μυελίνωσης, μερικές φορές είναι ορατές μικρές ρωγμές ή περιοχές του φυσιολογικού βυθού. Στο 17-20% των περιπτώσεων ανιχνεύονται αμφοτερόπλευρες μυελινωμένες νευρικές ίνες. Στο 19% των περιπτώσεων δεν έρχονται σε επαφή με την κεφαλή του οπτικού νεύρου. Περιστασιακά, μεμονωμένες δέσμες μυελινωμένων νευρικών ινών ανιχνεύονται στην περιφέρεια του αμφιβληστροειδούς στη ρινική πλευρά του ΟΝΗ.
Η παθογένεση των ινών μυελίνηςπαραμένει ασαφές, αλλά τα ζώα με αδύναμο ή απών lamina cribrosa έχουν συνήθως βαθιά φυσιολογική εκσκαφή και εκτεταμένη μυελίωση των νευρικών ινών του αμφιβληστροειδούς, ενώ τα ζώα με ένα καλά ανεπτυγμένο αδρανές έλασμα παρουσιάζουν εντελώς επίπεδο οπτικό δίσκο (OND) και έλλειψη μυελίνωσης του αμφιβληστροειδούς. προτείνει διάφορους πιθανούς μηχανισμούς παθογένεσης:
1. Η παρουσία ενός ελαττώματος στην αδρή πλάκα, που επιτρέπει στα ολιγοδενδροκύτταρα να διεισδύσουν στον αμφιβληστροειδή και να παράγουν μυελίνη.
2. Είναι πιθανό ο αριθμός των αξόνων να είναι μικρός σε σύγκριση με το μέγεθος του σκληρού σωλήνα και να υπάρχει αρκετός χώρος για να συνεχιστεί η μυελίνωση στην κοιλότητα του ματιού. Σε μάτια με περιφερικά απομονωμένες δέσμες μυελινωμένων νευρικών ινών, η διαταραχή του χρονισμού του λεπτού ελάσματος επιτρέπει στα ολιγοδενδροκύτταρα να εισέλθουν στον αμφιβληστροειδή και να μεταναστεύσουν στο στρώμα των νευρικών ινών μέχρι να φτάσουν σε μια περιοχή σχετικά χαμηλής πυκνότητας νευρικών ινών όπου συνεχίζεται η μυελίνωση.
3. Η όψιμη ανάπτυξη της αδρανούς πλάκας μπορεί να δημιουργήσει συνθήκες για τη μετανάστευση των ολιγοδενδροκυττάρων στους ιστούς του οφθαλμού.
Η εκτεταμένη μονόπλευρη (ή, σπάνια, αμφοτερόπλευρη) μυελίνωση των νευρικών ινών μπορεί να συνοδεύεται από υψηλή μυωπία και διαθλαστική αμβλυωπία. Σε αυτούς τους ασθενείς, η μυελίνη καλύπτει το μεγαλύτερο μέρος, αν όχι όλη, την περιφέρεια του δίσκου. Στην περιοχή της ωχράς κηλίδας (αν και απαλλαγμένη από μυελίνη), συνήθως ανιχνεύονται επίσης ανωμαλίες - εξασθένηση του αντανακλαστικού ή διασπορά της χρωστικής. Η κατάσταση της περιοχής της ωχράς κηλίδας είναι ίσως ο πιο ακριβής προγνωστικός παράγοντας για την αποτελεσματικότητα της αποφρακτικής θεραπείας.
μυελινωμένες νευρικές ίνεςμπορεί να συνοδεύεται από σύνδρομο Gorlin (πολλαπλοί βασικοκυτταρικοί σπίλοι) και αυτοσωμική κυρίαρχη υαλοαμφιβληστροειδοπάθεια, συνοδευόμενη από συγγενή οπτική βλάβη, αμφοτερόπλευρη εκτεταμένη μυελίωση της στιβάδας των νευρικών ινών του αμφιβληστροειδούς, σοβαρή εκφύλιση του υαλοειδούς, υψηλή μυωπία, δυστροφία αμφιβληστροειδούς, νυχτερινή τύφλωση και παραμορφώσεις άκρων.
μυελινωμένες νευρικές ίνεςμπορεί να κληρονομηθεί με αυτοσωμικό κυρίαρχο τρόπο. Έχουν περιγραφεί μεμονωμένες περιπτώσεις μυελινωμένων νευρικών ινών που σχετίζονται με ανώμαλο μακρύ οπτικό νεύρο (οξυκεφαλία), ελαττώματα στο κριβρόσωμα του ελάσματος (λοξός οπτικός δίσκος), δυσγένεση του πρόσθιου τμήματος και νευροϊνωμάτωση τύπου 2. Αν και οι μυελινωμένες νευρικές ίνες μπορεί να συνυπάρχουν με νευροϊνωμάτωση, πολλοί συγγραφείς θεωρούν αυτόν τον συνδυασμό αμφισβητήσιμο.
Περιστασιακά ζώνες μυελίνωση των νευρικών ινώνμπορεί να εμφανιστεί μετά τη βρεφική ηλικία και ακόμη και στην ενήλικη ζωή. Σε τέτοιες περιπτώσεις, το τραύμα στο μάτι (αμβλύ τραύμα στο μάτι σε μια περίπτωση και φούσκωμα των περιβλημάτων του οπτικού νεύρου σε μια άλλη) είναι πιθανώς η συνήθης αιτία αυτής της ανωμαλίας.
Είναι πιθανό ότι μια τέτοια ζημιά υπάρχει ένα ελάττωμα στην ακανθώδη πλάκαεπαρκής για τη διείσδυση ολιγοδενδροκυττάρων στον αμφιβληστροειδή. Οι μυελινωμένες νευρικές ίνες μπορεί να εξαφανιστούν όταν οι άξονες καταστραφούν.
Μονόπλευρη μυωπία υψηλού βαθμού με μυελινωμένες νευρικές ίνες. μυελινωμένες νευρικές ίνες.
μυελινωμένες νευρικές ίνες. Α. Ελαφρύ βαθμό. Β. Βαρύς βαθμός.
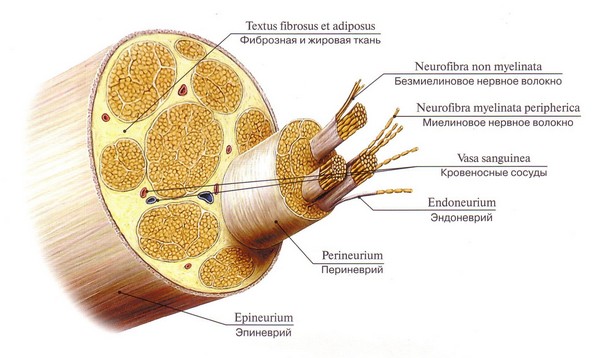
Οι διεργασίες των νευρώνων καλύπτονται σχεδόν πάντα με ένα περίβλημα (μυελίνη). Εξαίρεση αποτελούν οι ελεύθερες απολήξεις ορισμένων διεργασιών. Η διαδικασία μαζί με το έλυτρο ονομάζεται «νευρική ίνα».
Η νευρική ίνα αποτελείται από: κύλινδρος άξονα- ανάπτυξη νευρικού κυττάρου: άξονας ή δενδρίτης
Γλοιακό περίβλημα, που περιβάλλει τον αξονικό κύλινδρο σε μορφή ζεύξης. Στο ΚΝΣ, σχηματίζεται από ολιγοδενδρογλοία και στο ΠΝΣ από κύτταρα Schwann (τα νευρολεμοκύτταρα είναι ένας τύπος ολιγοδενδρογλοίας).
Οι νευρικές ίνες ταξινομούνται σε μη μυελινωμένες και μυελινωμένες (που έχουν θηκάρι μυελίνης).
Οι μη μυελινωμένες νευρικές ίνες αποτελούν μέρος του αυτόνομου νευρικού συστήματος και αντιπροσωπεύονται από άξονες τελεστικών νευρώνων. Βρίσκονται επίσης στο κεντρικό νευρικό σύστημα, αλλά σε μικρότερες ποσότητες.
Δομή: Στο κέντρο βρίσκεται ο πυρήνας ενός ολιγοδενδροκυττάρου (λεμοκύτταρο), και κατά μήκος της περιφέρειας 10-20 αξονικοί κύλινδροι διεισδύουν στο κυτταρόπλασμά του. Τέτοιες νευρικές ίνες ονομάζονται επίσης "ίνες τύπου καλωδίου". Όταν ο αξονικός κύλινδρος βυθίζεται στο κυτταρόπλασμα του ολιγοδενδροκυττάρου, τα τμήματα του πλασμολήμματος του τελευταίου πλησιάζουν το ένα το άλλο και σχηματίζεται ένα μεσεντέριο - "μεσάξον" ή διπλή μεμβράνη. Από την επιφάνεια, η νευρική ίνα καλύπτεται με μια βασική μεμβράνη.
Οι μυελινωμένες νευρικές ίνες αποτελούν μέρος του ΚΝΣ, των σωματικών υποδιαιρέσεων του ΠΝΣ και των προγαγγλιακών τμημάτων του αυτόνομου νευρικού συστήματος. Μπορεί να περιέχουν τόσο άξονες όσο και δενδρίτες νευρώνων.
Δομή: Ο αξονικός κύλινδρος είναι πάντα 1, βρίσκεται στο κέντρο. Το κέλυφος έχει 2 στρώματα: το εσωτερικό (μυελίνη) και το εξωτερικό (νευρολήμμα), που αντιπροσωπεύεται από τον πυρήνα και το κυτταρόπλασμα του κυττάρου Schwann. Εξωτερικά υπάρχει μια βασική μεμβράνη. Το στρώμα μυελίνης είναι πολλά στρώματα της μεμβράνης του ολιγοδενδροκυττάρου (λεμοκύτταρο). Η μεμβράνη είναι ομόκεντρα στριμμένη γύρω από τον αξονικό κύλινδρο. Στην πραγματικότητα, πρόκειται για ένα πολύ επιμήκη μεσαξόνιο. Τα μεσαξόνια σχηματίζουν γλωσσικές κυτταροπλασματικές διεργασίες.
Η διαδικασία της μυελίνωσης είναι ο σχηματισμός του περιβλήματος μυελίνης. Εμφανίζεται στα τελευταία στάδια της εμβρυογένεσης και τους πρώτους μήνες μετά τη γέννηση.
Αξίζει να σημειωθεί ότι υπάρχουν χαρακτηριστικά μυελίνωσης στο ΚΝΣ: 1 ολιγοδενδροκύτταρο σχηματίζει ένα περίβλημα μυελίνης γύρω από αρκετούς αξονικούς κυλίνδρους (με τη βοήθεια πολλών διεργασιών που περιστρέφονται). Δεν υπάρχει βασική μεμβράνη.
Η δομή της ίνας της μυελίνης.
Η μυελίνη διακόπτεται τακτικά στους κόμβους του Ranvier. Η απόσταση μεταξύ των παρεμβολών είναι 0,3 - 1,5 nm. Στην περιοχή της αναχαίτισης πραγματοποιείται ο τροφισμός του αξονικού κυλίνδρου. Η μυελίνη έχει εγκοπές στην επιφάνειά της. Αυτά τα τμήματα κοπής μυελίνης αυξάνουν την ευκαμψία της νευρικής ίνας και αποτελούν «απόθεμα» για τέντωμα. Δεν υπάρχουν εγκοπές στο ΚΝΣ.
Η μυελίνη χρωματίζεται με βαφές για λιπίδια: Σουδάν, οσμικό οξύ.
Λειτουργίες μυελίνης:
Αύξηση της ταχύτητας αγωγής των νευρικών παλμών. Οι ίνες χωρίς μυελίνη έχουν ταχύτητα 1-2 m/s, ενώ οι μυελινωμένες ίνες έχουν ταχύτητα 5-120 m/s.
Τα κανάλια Na είναι συγκεντρωμένα στην περιοχή των τομέων, όπου προκύπτουν βιοηλεκτρικά ρεύματα. Πηδάνε από τη μια αναχαίτιση στην άλλη. Αυτή είναι η αλμυρή αγωγή, δηλαδή η αγωγή μιας ώθησης στα άλματα.
Η μυελίνη είναι ένας μονωτήρας που περιορίζει την είσοδο των ρευμάτων που εξαπλώνονται γύρω.
Η διαφορά στη δομή των μυελινωμένων και μη μυελινωμένων ινών.
μη μυελιωμένη ίνα μυελινωμένη ίνα
Κύλινδροι πολλών αξόνων 1 κύλινδρος άξονα
Αξονικοί κύλινδροι - άξονες
Ο πυρήνας του ολιγοδενδροκυττάρου βρίσκεται στο κέντρο Ο πυρήνας και το κυτταρόπλασμα του ολιγοδενδροκυττάρου βρίσκονται στην περιφέρεια της ίνας
Οι μεσάξονες είναι κοντοί Οι μεσάξονες περιστρέφονται επανειλημμένα γύρω από τον αξονικό κύλινδρο, σχηματίζεται το περίβλημα της μυελίνης
Κανάλια Na σε όλο το μήκος του αξονικού κυλίνδρου κανάλια Na μόνο στους κόμβους του Ranvier
Η δομή του περιφερικού νεύρου.
Το νεύρο αποτελείται από μυελινωμένες και μη μυελινωμένες ίνες ομαδοποιημένες σε δεσμίδες. Περιέχει τόσο προσαγωγές όσο και απαγωγές ίνες.
Μηχανισμοί αγωγιμότητας νευρικών παλμών.
Οι συνάψεις είναι ειδικές διακυτταρικές συνδέσεις που χρησιμοποιούνται για τη μεταφορά ενός σήματος από το ένα κύτταρο στο άλλο.
Οι περιοχές επαφής των νευρώνων είναι πολύ κοντά μεταξύ τους. Ωστόσο, μεταξύ τους συχνά παραμένει ένα συναπτικό χάσμα που τους χωρίζει. Το πλάτος της συναπτικής σχισμής είναι της τάξης πολλών δεκάδων νανομέτρων.
Για να λειτουργήσουν επιτυχώς τα νετρόνια, είναι απαραίτητο να διασφαλιστεί η απομόνωσή τους μεταξύ τους και η αλληλεπίδραση μεταξύ τους παρέχεται από τις συνάψεις.
Οι συνάψεις λειτουργούν ως ενισχυτές των νευρικών σημάτων κατά μήκος της διαδρομής τους. Το αποτέλεσμα επιτυγχάνεται από το γεγονός ότι μια σχετικά χαμηλής ισχύος ηλεκτρική ώθηση απελευθερώνει εκατοντάδες χιλιάδες μόρια μεσολαβητών, τα οποία περιέχονταν προηγουμένως σε πολλά συναπτικά κυστίδια. Ένα βολίδα μορίων μεσολαβητών δρα συγχρονισμένα σε μια μικρή περιοχή του ελεγχόμενου νευρώνα, όπου συγκεντρώνονται μετασυναπτικοί υποδοχείς - εξειδικευμένες πρωτεΐνες που μετατρέπουν το σήμα τώρα από χημική μορφή σε ηλεκτρική.
Επί του παρόντος, τα κύρια στάδια της διαδικασίας απελευθέρωσης του μεσολαβητή είναι καλά γνωστά. Μια νευρική ώθηση, δηλαδή ένα ηλεκτρικό σήμα, προκύπτει σε έναν νευρώνα, διαδίδεται κατά μήκος των διεργασιών του και φτάνει στις νευρικές απολήξεις. Η μετατροπή του σε χημική μορφή ξεκινά με το άνοιγμα διαύλων ιόντων ασβεστίου στην προσυναπτική μεμβράνη, η κατάσταση των οποίων ελέγχεται από το ηλεκτρικό πεδίο της μεμβράνης. Τώρα τα ιόντα ασβεστίου αναλαμβάνουν το ρόλο των φορέων σήματος. Εισέρχονται από τα ανοιχτά κανάλια μέσα στη νευρική απόληξη. Μια απότομα αυξημένη συγκέντρωση ιόντων ασβεστίου κοντά στη μεμβράνη για σύντομο χρονικό διάστημα ενεργοποιεί τη μοριακή μηχανή για την απελευθέρωση μεσολαβητή: τα συναπτικά κυστίδια πηγαίνουν στα σημεία της επακόλουθης σύντηξής τους με την εξωτερική μεμβράνη και, τέλος, ρίχνουν το περιεχόμενό τους στο χώρο της συναπτικής σχισμής .
Η συναπτική μετάδοση πραγματοποιείται με μια ακολουθία δύο χωρικά διαχωρισμένων διεργασιών: προσυναπτική στη μία πλευρά του συναπτικού κενού και μετασυναπτική στην άλλη (Εικ. 3). Οι απολήξεις των διεργασιών του νευρώνα ελέγχου, υπακούοντας στα ηλεκτρικά σήματα που έχουν έρθει σε αυτά, απελευθερώνουν μια ειδική ενδιάμεση ουσία (μεσολαβητή) στο χώρο της συναπτικής σχισμής. Τα μόρια του μεσολαβητή διαχέονται μάλλον γρήγορα μέσω της συναπτικής σχισμής και διεγείρουν ένα ηλεκτρικό σήμα απόκρισης στο ελεγχόμενο κύτταρο (άλλος νευρώνας, μυϊκή ίνα, ορισμένα κύτταρα των εσωτερικών οργάνων). Περίπου δώδεκα διαφορετικές ουσίες χαμηλού μοριακού βάρους λειτουργούν ως μεσολαβητές:
ακετυλοχολίνη (ένας εστέρας της αμινοαλκοόλης της χολίνης και του οξικού οξέος), γλουταμινικό (ανιόν του γλουταμινικού οξέος), GABA (γάμα-αμινοβουτυρικό οξύ), σεροτονίνη (ένα παράγωγο του αμινοξέος τρυπτοφάνη), αδενοσίνη, κ.λπ.
Συντίθενται προκαταρκτικά από τον προσυναπτικό νευρώνα από διαθέσιμες και σχετικά φθηνές πρώτες ύλες και αποθηκεύονται μέχρι να χρησιμοποιηθούν σε συναπτικά κυστίδια, όπου, σαν σε δοχεία, περικλείονται πανομοιότυπα τμήματα του μεσολαβητή (πολλές χιλιάδες μόρια σε ένα κυστίδιο).
διάγραμμα συνάψεων
Πάνω - ένα τμήμα της νευρικής απόληξης, που περιορίζεται από την προσυναπτική μεμβράνη, στην οποία είναι ενσωματωμένοι οι προσυναπτικοί υποδοχείς. τα συναπτικά κυστίδια μέσα στην νευρική απόληξη είναι γεμάτα με μεσολαβητή και είναι σε διάφορους βαθμούς ετοιμότητας για την απελευθέρωσή του. οι μεμβράνες των κυστιδίων και η προσυναπτική μεμβράνη περιέχουν προσυναπτικές πρωτεΐνες. Παρακάτω - ένα τμήμα ενός ελεγχόμενου κυττάρου, στην μετασυναπτική μεμβράνη του οποίου είναι ενσωματωμένοι μετασυναπτικοί υποδοχείς.
Οι συνάψεις είναι ένα βολικό αντικείμενο για τη ρύθμιση των ροών πληροφοριών. Το επίπεδο ενίσχυσης του σήματος κατά τη μετάδοσή του μέσω της σύναψης μπορεί εύκολα να αυξηθεί ή να μειωθεί αλλάζοντας την ποσότητα του μεσολαβητή που απελευθερώνεται, μέχρι την πλήρη απαγόρευση της μετάδοσης πληροφοριών. Θεωρητικά, αυτό μπορεί να γίνει στοχεύοντας οποιοδήποτε από τα στάδια απελευθέρωσης μεσολαβητή.
Ίνες μυελίνης. Μέρος των νευρικών ινών κατά τη διάρκεια της εμβρυογένεσης υφίσταται μυελίωση: τα λεμοκύτταρα (κύτταρα Schwann) πρώτα αγγίζουν τον άξονα και μετά τον περιβάλλουν. Η μεμβράνη των λεμοκυττάρων τυλίγεται γύρω από τον άξονα σαν ρολό, σχηματίζοντας μια πολυστρωματική έλικα (θηκάρι μυελίνης). Το περίβλημα μυελίνης δεν είναι συνεχές - σε όλο το μήκος της νευρικής ίνας σε ίση απόσταση το ένα από το άλλο, υπάρχουν μικρά σπασίματα σε αυτό (ο Ranvier παρεμποδίζει). Στην περιοχή των τομών, ο άξονας στερείται θήκης μυελίνης.
μη μυελινωμένες ίνες.Η μυελίνωση των άλλων ινών τελειώνει στα αρχικά στάδια της εμβρυϊκής ανάπτυξης. Ένας ή περισσότεροι άξονες βυθίζονται στο λεμοκύτταρο. τα περιβάλλει πλήρως ή εν μέρει, αλλά δεν σχηματίζει πολυστρωματικό περίβλημα μυελίνης.
Ο μηχανισμός αγωγής της διέγερσης κατά μήκος μη μυελινωμένων νευρικών ινών
Σε ηρεμία, ολόκληρη η εσωτερική επιφάνεια της μεμβράνης της νευρικής ίνας φέρει αρνητικό φορτίο και η εξωτερική πλευρά της μεμβράνης είναι θετική. Το ηλεκτρικό ρεύμα μεταξύ της εσωτερικής και της εξωτερικής πλευράς της μεμβράνης δεν ρέει, επειδή. η λιπιδική μεμβράνη έχει υψηλή ηλεκτρική αντίσταση.Κατά την ανάπτυξη του δυναμικού δράσης στην διεγερμένη περιοχή της μεμβράνης, εμφανίζεται μια αναστροφή φορτίου. Στα όρια της διεγερμένης και μη διεγερμένης περιοχής, αρχίζει να ρέει ένα ηλεκτρικό ρεύμα. Ένα ηλεκτρικό ρεύμα ερεθίζει το πλησιέστερο τμήμα της μεμβράνης και το φέρνει σε κατάσταση διέγερσης, ενώ τα προηγουμένως διεγερμένα τμήματα επιστρέφουν σε κατάσταση ηρεμίας. Έτσι, το κύμα διέγερσης καλύπτει όλα τα νέα τμήματα της μεμβράνης των νευρικών ινών.
Ο μηχανισμός αγωγής της διέγερσης κατά μήκος των νευρικών ινών της μυελίνης
Σε μια μυελινωμένη νευρική ίνα, τα τμήματα της μεμβράνης που καλύπτονται από το περίβλημα της μυελίνης είναι μη διεγέρσιμα. Η διέγερση μπορεί να συμβεί μόνο σε περιοχές της μεμβράνης που βρίσκονται στην περιοχή των κόμβων του Ranvier. Με την ανάπτυξη του AP σε έναν από τους κόμβους του Ranvier, το φορτίο της μεμβράνης αντιστρέφεται. Ένα ηλεκτρικό ρεύμα προκύπτει ανάμεσα στα ηλεκτραρνητικά και ηλεκτροθετικά τμήματα της μεμβράνης, το οποίο ερεθίζει γειτονικά τμήματα της μεμβράνης. Ωστόσο, μόνο ένα τμήμα της μεμβράνης στην περιοχή του επόμενου κόμβου του Ranvier μπορεί να εισέλθει στην κατάσταση διέγερσης. Έτσι, η διέγερση εξαπλώνεται σε όλη τη μεμβράνη με αλματώδη (αλατώδη) τρόπο από τη μια αναχαίτιση του Ranvier στην άλλη.
ΛΕΙΤΟΥΡΓΙΕΣ ΕΝΟΣ ΝΕΥΡΩΝΑ
Το υλικό για την κατασκευή του κεντρικού νευρικού συστήματος και των αγωγών του είναι ο νευρικός ιστός, ο οποίος αποτελείται από δύο συστατικά - νευρικά κύτταρα (νευρώνες) και νευρογλοία. Τα κύρια λειτουργικά στοιχεία του ΚΝΣ είναι οι νευρώνες: στο σώμα των ζώων υπάρχουν περίπου 50 δισεκατομμύρια από αυτά, από τα οποία μόνο ένα μικρό μέρος βρίσκεται στα περιφερειακά μέρη του σώματος.
Οι νευρώνες αποτελούν το 10 - 15% του συνολικού αριθμού των κυτταρικών στοιχείων στο νευρικό σύστημα. Το κύριο μέρος του καταλαμβάνεται από νευρογλοιακά κύτταρα. Στα ανώτερα ζώα, στη διαδικασία της μεταγεννητικής οντογένεσης, οι διαφοροποιημένοι νευρώνες δεν διαιρούνται. (όπως πιστεύαμε προηγουμένως). Οι νευρώνες διαφέρουν σημαντικά ως προς το σχήμα (πυραμιδικό, στρογγυλό, αστερικό, οβάλ), το μέγεθος (από 5 έως 150 μικρά), τον αριθμό των διεργασιών, αλλά έχουν επίσης κοινές ιδιότητες.
Σε ορισμένα μέρη του εγκεφάλου υπάρχουν νευρώνες που παράγουν κόκκους έκκρισης βλεννοπρωτεϊνικής ή γλυκοπρωτεϊνικής φύσης. Έχουν τόσο φυσιολογικά χαρακτηριστικά νευρώνων όσο και αδενικών κυττάρων. Αυτά τα κύτταρα ονομάζονται νευροεκκριτικά. Η λειτουργία των νευρώνων είναι να αντιλαμβάνονται τα σήματα από υποδοχείς ή άλλα νευρικά κύτταρα, να αποθηκεύουν και να επεξεργάζονται πληροφορίες και να μεταδίδουν νευρικές ώσεις σε άλλα κύτταρα - νευρικά, μυϊκά ή εκκριτικά κύτταρα. Αντίστοιχα, υπάρχει μια εξειδίκευση των νευρώνων. Χωρίζονται σε 3 ομάδες: ευαίσθητους (αισθητηριακούς, προσαγωγούς) νευρώνες, συνειρμικούς (ενδιάμεσους, ενδιάμεσους) νευρώνες, κινητικούς (ενεργητικούς) νευρώνες.
Νευρογλοία. Ο κύριος όγκος του νευρικού ιστού αποτελείται από γλοιακά στοιχεία που εκτελούν βοηθητικές λειτουργίες και γεμίζουν σχεδόν ολόκληρο το διάστημα μεταξύ των νευρώνων. Ανατομικά, τα νευρογλοιακά κύτταρα στον εγκέφαλο (ολιγοδενδροκύτταρα και αστροκύτταρα) και τα κύτταρα Schwann στο περιφερικό νευρικό σύστημα διακρίνονται μεταξύ τους. Τα ολιγοδενδροκύτταρα και τα κύτταρα Schwann σχηματίζουν περιβλήματα μυελίνης γύρω από τους άξονες. Μεταξύ των νευρογλοιακών κυττάρων και των νευρώνων υπάρχουν κενά πλάτους 15 - 20 nm, τα οποία επικοινωνούν μεταξύ τους, σχηματίζοντας έναν διάμεσο χώρο γεμάτο με υγρό. Μέσω αυτού του χώρου, υπάρχει ανταλλαγή ουσιών μεταξύ του νευρώνα και των νευρογλοιακών κυττάρων, καθώς και η παροχή οξυγόνου και θρεπτικών ουσιών στους νευρώνες με διάχυση. Τα νευρογλοιακά κύτταρα, προφανώς, εκτελούν μόνο υποστηρικτικές και προστατευτικές λειτουργίες στο ΚΝΣ και δεν είναι, όπως αναμενόταν, η πηγή διατροφής τους ή οι θεματοφύλακες πληροφοριών.
Τα νευρογλοιακά κύτταρα διαφέρουν από τους νευρώνες στις ιδιότητες της μεμβράνης: αντιδρούν παθητικά στο ηλεκτρικό ρεύμα, οι μεμβράνες τους δεν παράγουν ώθηση διάδοσης. Υπάρχουν σφιχτές συνδέσεις (περιοχές χαμηλής αντίστασης) μεταξύ των νευρογλοιακών κυττάρων που παρέχουν άμεση ηλεκτρική σύνδεση. Το δυναμικό της μεμβράνης των νευρογλοιακών κυττάρων είναι υψηλότερο από αυτό των νευρώνων και εξαρτάται κυρίως από τη συγκέντρωση των ιόντων Κ+ στο μέσο.
Όταν η συγκέντρωση του Κ+ αυξάνεται κατά την ενεργό δραστηριότητα των νευρώνων στον εξωκυττάριο χώρο, ένα μέρος του απορροφάται από αποπολωμένα γλοιακά στοιχεία. Αυτή η ρυθμιστική λειτουργία της γλοίας εξασφαλίζει μια σχετικά σταθερή εξωκυτταρική συγκέντρωση Κ+. Τα γλοιακά κύτταρα - αστροκύτταρα - βρίσκονται μεταξύ των σωμάτων των νευρώνων και του τοιχώματος των τριχοειδών αγγείων, οι διεργασίες τους έρχονται σε επαφή με το τοίχωμα των τελευταίων. Αυτές οι περιαγγειακές διεργασίες είναι στοιχεία του αιματοεγκεφαλικού φραγμού. Τα μικρογλοιακά κύτταρα εκτελούν μια φαγοκυτταρική λειτουργία, ο αριθμός τους αυξάνεται απότομα όταν ο εγκεφαλικός ιστός έχει υποστεί βλάβη.
Φυσιολογία συνάψεων.
Χημικές συνάψειςέχουν σχετικά ευρεία συναπτική σχισμή (20 - 50 nm) και υψηλή αντοχή των συναπτικών μεμβρανών. Το άκρο του προσυναπτικού νεύρου περιέχει μεγάλο αριθμό κυστιδίων - συναπτικά κυστίδια - περίπου 50 nm σε διάμετρο, γεμάτο με μεσολαβητή.
Ο μηχανισμός μετάδοσης διέγερσης σε χημικές διεγερτικές συνάψεις. Σε συνάψεις με χημική μετάδοση, η διέγερση μεταδίδεται χρησιμοποιώντας διαμεσολαβητές(μεσάζοντες). Διαμεσολαβητής ορία -Αυτές είναι χημικές ουσίες που παρέχουν τη μετάδοση της διέγερσης στις συνάψεις. Οι διαμεσολαβητές, ανάλογα με τη φύση τους, χωρίζονται σε διάφορες ομάδες:
μονοαμίνες(ακετυλοχολίνη, ντοπαμίνη, νορεπινεφρίνη, σεροτονίνη, κ.λπ.);
Αμινοξέα(γάμα-αμινοβουτυρικό οξύ - GABA, γλουταμικό οξύ, γλυκίνη κ.λπ.)
νευροπεπτίδια(ουσία P, ενδορφίνες, νευροτενσίνη, ACTH, αγγειοτενσίνη, βαζοπρεσσίνη, σωματοστατίνη κ.λπ.). Ο μεσολαβητής σε μοριακή μορφή βρίσκεται στα κυστίδια της προσυναπτικής πάχυνσης (συναπτική πλάκα), όπου εισέρχεται:
Από την περιπυρηνική περιοχή του νευρώνα χρησιμοποιώντας γρήγορη αξονική μεταφορά (αξονικό ρεύμα).
Λόγω της σύνθεσης ενός μεσολαβητή που εμφανίζεται στα συναπτικά τερματικά από τα προϊόντα διάσπασής του.
Λόγω της επανασύλληψης του νευροδιαβιβαστή από τη συναπτική σχισμή σε αμετάβλητη μορφή.
Ποικιλία συστημάτων διαμεσολαβητών. Αρχή Dale: ένας νευρώνας, κατά κανόνα, συνθέτει και χρησιμοποιεί έναν μεσολαβητή σε όλα τα τερματικά του. Είναι δυνατό για έναν νευρώνα να χρησιμοποιήσει αρκετούς μεσολαβητές (κωμικοί) , αλλά προφανώς στον ίδιο συνδυασμό (αυτή η αρχή αμφισβητείται επί του παρόντος, βλέπε παρακάτω για συστήματα διαμεσολαβητών) . Ως συνέπεια της αρχής Dale, μπορούν να ληφθούν υπόψη οι ακόλουθες διατάξεις:
1) Το σημάδι της συναπτικής δράσης καθορίζεται όχι από τον μεσολαβητή, αλλά από τις ιδιότητες των υποδοχέων στο μετασυναπτικό κύτταρο.
2) Οι υποδοχείς σε κύτταρα που είναι μετασυναπτικοί σε έναν μόνο προσυναπτικό νευρώνα μπορεί να διαφέρουν φαρμακολογικά και μπορεί να ελέγχουν διαφορετικούς διαύλους ιόντων.
3) Ένα μετασυναπτικό κύτταρο μπορεί να έχει περισσότερους από έναν τύπους υποδοχέων για έναν δεδομένο μεσολαβητή και καθένας από αυτούς τους υποδοχείς μπορεί να ελέγχει έναν διαφορετικό μηχανισμό ιοντικής αγωγής.
Λόγω αυτών των τριών ιδιοτήτων, τα κύτταρα μπορούν να ασκούν αντίθετες συναπτικές δράσεις τόσο σε διαφορετικά μετασυναπτικά κύτταρα όσο και στο ίδιο.
Οι μεσολαβητές που έχουν εντοπιστεί μέχρι σήμερα σε ζώα και ανθρώπους αποτελούν μια μάλλον ετερογενή ομάδα ουσιών. Μονοαμίνες: ακετυλοχολίνη, ντοπαμίνη, νορεπινεφρίνη, σεροτονίνη (5-υδροξυτρυπταμίνη, 5-ΗΤ), ισταμίνη. Αμινοξέα: γ-αμινοβουτυρικό οξύ (GABA), γλουταμινικό οξύ, γλυκίνη, ταυρίνη κ.λπ. ,βαζοπρεσίνη και τα λοιπά . Οι αντίστοιχες συνάψεις ονομάζονται, για παράδειγμα, χολινεργικά, σεροτονινεργικά, νοραδρενεργικό και τα λοιπά. Ένας μεγάλος αριθμός μεσολαβητών αντιστοιχεί σε μεγάλο αριθμό μετασυναπτικών υποδοχέων: χολινεργικούς υποδοχείς , αδρενοϋποδοχείς, υποδοχείς GABA και τα λοιπά. Μέσα σε κάθε ομάδα υποδοχέων, υπάρχει μια ποικιλία από τους υποτύπους τους, για παράδειγμα, νικοτινικοί και μουσκαρινικοί υποδοχείς ακετυλοχολίνης (H-XR και M-XR, αντίστοιχα).
Η διαίρεση των υποδοχέων πραγματοποιείται με βάση τις διαφορές στις φαρμακολογικές τους ιδιότητες: διαφορετικοί αγωνιστές (ουσίες που μιμούνται την επίδραση ενός μεσολαβητή) και ανταγωνιστές (ουσίες που εμποδίζουν την εκδήλωση της επίδρασης του μεσολαβητή). Για παράδειγμα, για το N-ChR, ο αγωνιστής είναι η νικοτίνη , ανταγωνιστές - τουβοκουραρίνη (απομονωμένη από το δηλητήριο του curare), μπουνγκαροτοξίνη (απομονωμένη από το δηλητήριο ενός φιδιού του γένους Bungarus). Υπάρχουν τρεις τύποι υποδοχέων γλουταμικού που βασίζονται σε αγωνιστές: quisqualate (τύπου AMPA), kainate Και NMDA (Ν-μεθυλ-D-ασπαρτικός) -τύπος. Ορισμένοι υποδοχείς μεσολαβητών (ιδιαίτερα, αδρενεργικοί υποδοχείς και υποδοχείς για πολλά νευροπεπτίδια) δεν σχετίζονται με διαύλους ιόντων (ιονοτροπικοί υποδοχείς) , αλλά με ένα ένζυμο μεμβράνης (μεταβοτροπικοί υποδοχείς) , για παράδειγμα, αδενυλική κυκλάση . Το τελευταίο, μόλις ενεργοποιηθεί από έναν μεσολαβητή, καταλύει τον μετασχηματισμό πολλών μορίων τριφωσφορική αδενοσίνη (ATP) κυκλικός μονοφωσφορική αδενοσίνη (cAMP) - μηχανισμός ενίσχυσης. Το CAMP, ως δεύτερος αγγελιοφόρος, ενεργοποιεί πολλά ένζυμα στο κύτταρο, ιδιαίτερα πρωτεϊνικές κινάσες και έτσι διεγείρει τον κυτταρικό μεταβολισμό. Το cAMP καταστρέφεται φωσφοδιεστεράση. Το σύστημα αδενυλικής κυκλάσης είναι παρόμοιο με το σύστημα γουανυλικής κυκλάσης, το σύστημα φωσφολιπάσης C κ.λπ. (Εικ. 14). Ο σχηματισμός cGMP από τη μη κυκλική μορφή καταλύεται από γουανυλική κυκλάση , η δραστηριότητα του οποίου διεγείρεται από το μονοξείδιο του αζώτου. Τα μόρια της τελευταίας σχηματίζονται κατά την απαμίνωση της αργινίνης και το σχηματισμό κιτρουλίνης υπό τη δράση της συνθάση νιτρικού οξειδίου. Η δραστηριότητα αυτού του ενζύμου, με τη σειρά του, ρυθμίζεται από το σύμπλοκο Ca 2+ -καλμοδουλίνη. Έτσι, για παράδειγμα, οι υποδοχείς γλουταμικού, οι οποίοι πυροδοτούν την εισερχόμενη ροή ιόντων Ca 2+, ελέγχουν τις διακυμάνσεις στη συγκέντρωση του cGMP στο κυτταρόπλασμα των νευρώνων. Στον εγκέφαλο των σπονδυλωτών, η δραστηριότητα της συνθάσης του μονοξειδίου του αζώτου βρίσκεται στην παρεγκεφαλίδα, στο τετράπλευρο, στο ραβδωτό σώμα και στον οσφρητικό βολβό.
Ο μηχανισμός της χημικής σύναψης:κατά την εκπόλωση του προσυναπτικού τερματικού (που προκαλείται από PD ή τεχνητά), ιόντα Ca 2+ εισέρχονται σε αυτό από το περιβάλλον, τα οποία διεγείρουν τη διαδικασία εξωκυττάρωση - άδειασμα κυστιδίων στη συναπτική σχισμή.
Παρουσιάζονται χαρακτηριστικές συναπτικές και φυσαλιδώδεις πρωτεΐνες μαζί με τους υποτιθέμενους υποδοχείς και τις λειτουργίες τους. Υποτίθεται ότι χωριστά τμήματα της φυσαλιδώδους μεμβράνης αγκυρώνουν τα κυστίδια στον κυτταροσκελετό), συνδέουν τη φυσαλιδώδη μεμβράνη στην προσυναπτική μεμβράνη και απελευθερώνουν τον μεσολαβητή μέσω του προκύπτοντος πόρου. Οι μοριακοί μηχανισμοί προσκόλλησης κυστιδίων στην προσυναπτική μεμβράνη και σχηματισμός πόρων είναι πιθανώς διαφορετικοί. Μερικές από τις πρωτεΐνες είναι στόχοι νευροτοξινών που αλλάζουν την απελευθέρωση του νευροδιαβιβαστή. Για παράδειγμα, η δομή των φυσαλιδωδών πρωτεϊνών συναπτομπρεβίνης (VAMPs) διαταράσσεται από τον τέτανο και τις τοξίνες αλλαντίασης. Το δηλητήριο της αράχνης, η λατροτοξίνη, συνδέεται με τις πρωτεΐνες της προσυναπτικής μεμβράνης, τις νευρεξίνες και ενισχύει την κένωση των κυστιδίων.
1. Οι συναψίνες είναι πρωτεΐνες που σχετίζονται με κυστίδια που πιθανώς συνδέουν τα συναπτικά κυστίδια με τον κυτταροσκελετό της νευρικής απόληξης. 2. Η προσκόλληση, ο σχηματισμός πόρων και η εκκένωση των κυστιδίων διαμεσολαβούνται από αλληλεπιδράσεις (που υποδεικνύονται με βέλη) διαφόρων κυστιδίων και μεμβρανικών πρωτεϊνών. Για παράδειγμα, οι φυσαλιδώδεις πρωτεΐνες (συναπτοταγμίνη και συναπτομπρεβίνες) και οι πρωτεΐνες της πλασματικής μεμβράνης της νευρικής απόληξης (συνταξίνες και νευρεξίνες) εμπλέκονται στο σχηματισμό του συμπλέγματος προσκόλλησης. 3. Ποιες πρωτεΐνες - πλασματική ή φυσαλιδώδης σχηματίζουν τον πόρο σύντηξης δεν είναι ακόμα σαφές. Πιθανώς, αυτές είναι η συναπτοφυσίνη (που έχει άλλες λειτουργίες) και η πρωτεΐνη της μεμβράνης του πλάσματος φυσοφιλίνη. 4. Οι πρωτεΐνες Rab μπορούν να συμμετέχουν στη μεταφορά κυστιδίων στο κύτταρο και στην προσκόλλησή τους στη μεμβράνη των νευρικών κυττάρων. Οι φυσαλιδώδεις μεταφορείς παίζουν ρόλο στη συσσώρευση νευροδιαβιβαστών στα συναπτικά κυστίδια.
Ταυτόχρονα, το Ca 2+ αρχίζει να απομακρύνεται από το κυτταρόπλασμα με διάφορους τρόπους: δέσμευση πρωτεϊνών, μιτοχονδριακή πρόσληψη και ενεργή μεταφορά. Η έξοδος του μεσολαβητή εξαρτάται από την εκπόλωση του τερματικού και είναι περίπου 100 - 200 κυστίδια, καθένα από τα οποία περιέχει ένα τμήμα (κβαντικό), που αντιστοιχεί σε περίπου 10 4 μόρια. Τα μόρια του μεσολαβητή διαχέονται στη μετασυναπτική μεμβράνη, όπου αλληλεπιδρά με τους υποδοχείς της μετασυναπτικής μεμβράνης που ρυθμίζουν την κατάσταση των διαύλων ιόντων. Αυτή η ρύθμιση μπορεί να είναι άμεση (όπως, για παράδειγμα, στη νευρομυϊκή σύνδεση των σκελετικών μυών των σπονδυλωτών) ή να περιλαμβάνει την ενεργοποίηση συστημάτων δευτερεύων ενδοκυτταρικοί μεσολαβητές (G πρωτεΐνες, cAMP).
Στην πρώτη περίπτωση, μεταδίδονται σήματα γρήγορης εκκίνησης, στη δεύτερη, πραγματοποιούνται πιο αργά μακροπρόθεσμα αποτελέσματα. Η κατεύθυνση της αλλαγής του δυναμικού της μετασυναπτικής μεμβράνης (αποπόλωση ή υπερπόλωση) εξαρτάται κυρίως από το άνοιγμα των καναλιών που ελέγχουν οι μετασυναπτικοί υποδοχείς. Μερικά από τα μόρια του μεσολαβητή μπορούν να αλληλεπιδράσουν προσυναπτικούς υποδοχείς, η οποία οδηγεί σε αλλαγή του MP του νευρικού τερματικού και, κατά συνέπεια, της ποσότητας του μεσολαβητή που απελευθερώνεται (ανάδραση). Η συναπτική σχισμή καθαρίζεται από τον μεσολαβητή με διάφορους τρόπους: απενεργοποίηση, υδρόλυση, επαναπρόσληψη στην προσυναπτική απόληξη, διάχυση, σύλληψη από γλοιακά κύτταρα. Κύριο μέρος συναπτική καθυστέρηση - ο χρόνος από την άφιξη μιας νευρικής ώθησης έως την ανάπτυξη μιας μετασυναπτικής απόκρισης (0,2-0,5 ms) πέφτει στη διαδικασία έκκρισης νευροδιαβιβαστών. Η χημική σύναψη παρέχει μετάδοση σήματος μόνο από τον προσυναπτικό νευρώνα στον μετασυναπτικό.
Με συχνή ρυθμική διέγερση σε χημικές συνάψεις, αύξηση σε (ανακούφιση) και μετά εξασθένιση (κατάθλιψη) μετάδοση, δηλ. αυξάνονται και στη συνέχεια πέφτουν στο πλάτος των μετασυναπτικών δυναμικών. Αυτά τα φαινόμενα καθορίζονται κυρίως από αλλαγές στον προσυναπτικό σύνδεσμο. Έχουν ιδιαίτερη ανάπτυξη σε ορισμένες συνάψεις του ΚΝΣ, όπου δρουν ως παράγοντες συναπτικής πλαστικότητας. Η παρουσία διαφόρων τύπων υποδοχέων στη μετασυναπτική μεμβράνη μπορεί να προκαλέσει την ανάπτυξη αυτών των φαινομένων ξεχωριστά, όπως η μακροχρόνια ενίσχυση και η μακροχρόνια κατάθλιψη.
Χημικές ανασταλτικές συνάψεις. Αυτές οι συνάψεις, ως προς τον μηχανισμό μετάδοσης της διέγερσης, είναι παρόμοιες με τις συνάψεις διεγερτικής δράσης. Σε ανασταλτικές συνάψεις, ένας μεσολαβητής (για παράδειγμα, γλυκίνη) αλληλεπιδρά με υποδοχείς στην υποσυναπτική μεμβράνη και ανοίγει κανάλια χλωρίου σε αυτήν, γεγονός που οδηγεί στην κίνηση ιόντων χλωρίου κατά μήκος της βαθμίδας συγκέντρωσης στο κύτταρο και στην ανάπτυξη υπερπόλωσης στην υποσυναπτική μεμβράνη . Υπάρχει ένα λεγόμενο ανασταλτικό μετασυναπτικό δυναμικό(TPSP).
Προηγουμένως, πιστευόταν ότι κάθε μεσολαβητής αντιστοιχεί σε μια συγκεκριμένη αντίδραση του μετασυναπτικού κυττάρου - διέγερση ή αναστολή με τη μία ή την άλλη μορφή. Έχει πλέον αποδειχθεί ότι ένας μεσολαβητής τις περισσότερες φορές αντιστοιχεί σε όχι έναν, αλλά σε πολλούς διαφορετικούς υποδοχείς. Για παράδειγμα, η ακετυλοχολίνη στις νευρομυϊκές συνάψεις των σκελετικών μυών δρα στους Η-χολινεργικούς υποδοχείς (ευαίσθητους στη νικοτίνη), οι οποίοι ανοίγουν μεγάλα κανάλια για νάτριο (και κάλιο), το οποίο δημιουργεί EPSP (PKP). Μ-χολινεργικοί υποδοχείς (ευαίσθητοι στη μουσκαρίνη), οι οποίοι ανοίγουν εκλεκτικούς διαύλους για ιόντα καλίου, έτσι δημιουργείται εδώ ανασταλτικό μετασυναπτικό δυναμικό (IPSP). Κατά συνέπεια, η διεγερτική ή ανασταλτική φύση της δράσης του μεσολαβητή καθορίζεται από τις ιδιότητες της υποσυναπτικής μεμβράνης (ακριβέστερα, από τον τύπο του υποδοχέα) και όχι από τον ίδιο τον μεσολαβητή.
Φυσιολογικές ιδιότητες χημικών συνάψεων. Οι συνάψεις με χημική μετάδοση της διέγερσης έχουν μια σειρά από κοινές ιδιότητες:
Η διέγερση μέσω των συνάψεων πραγματοποιείται μόνο προς μία κατεύθυνση (μονομερώς). Αυτό οφείλεται στη δομή της σύναψης: ο μεσολαβητής απελευθερώνεται μόνο από την προσυναπτική πάχυνση και αλληλεπιδρά με τους υποδοχείς της υποσυναπτικής μεμβράνης.
Η μετάδοση της διέγερσης μέσω των συνάψεων είναι πιο αργή από ότι μέσω της νευρικής ίνας - συναπτική καθυστέρηση.
Η μεταφορά της διέγερσης πραγματοποιείται με τη βοήθεια ειδικών χημικών μεσολαβητών - μεσολαβητών.
Στις συνάψεις, ο ρυθμός της διέγερσης μετασχηματίζεται.
Οι συνάψεις έχουν χαμηλή αστάθεια.
Οι συνάψεις είναι πολύ κουρασμένες.
Οι συνάψεις είναι ιδιαίτερα ευαίσθητες σε χημικές (συμπεριλαμβανομένων των φαρμακολογικών) ουσιών.
Διεγερτικές ηλεκτρικές συνάψεις. Εκτός από τις συνάψεις με χημική μετάδοση της διέγερσης, οι συνάψεις με ηλεκτρική μετάδοση βρίσκονται κυρίως στο κεντρικό νευρικό σύστημα (ΚΝΣ). Οι διεγερτικές ηλεκτρικές συνάψεις χαρακτηρίζονται από μια πολύ στενή συναπτική σχισμή και μια πολύ χαμηλή ειδική αντίσταση γειτονικών προ- και μετασυναπτικών μεμβρανών, η οποία εξασφαλίζει την αποτελεσματική διέλευση των τοπικών ηλεκτρικών ρευμάτων. Η χαμηλή αντίσταση, κατά κανόνα, συνδέεται με την παρουσία εγκάρσιων καναλιών που διασχίζουν και τις δύο μεμβράνες, δηλ. πηγαίνουν από κύτταρο σε κύτταρο (επαφή με διάκενο). Τα κανάλια σχηματίζονται από πρωτεϊνικά μόρια (ημιμόρια) καθεμιάς από τις μεμβράνες επαφής, τα οποία συνδέονται συμπληρωματικά. Αυτή η δομή είναι εύκολα βατή για ηλεκτρικό ρεύμα.
Διάγραμμα μετάδοσης διέγερσης σε ηλεκτρική σύναψη: το ρεύμα που προκαλείται από το προσυναπτικό δυναμικό δράσης ερεθίζει τη μετασυναπτική μεμβράνη, όπου εμφανίζονται το EPSP και το δυναμικό δράσης. Τα εγκάρσια κανάλια ενώνουν τα κύτταρα όχι μόνο ηλεκτρικά, αλλά και χημικά, καθώς είναι βατά για πολλές ενώσεις χαμηλού μοριακού βάρους. Επομένως, διεγερτικές ηλεκτρικές συνάψεις με εγκάρσια κανάλια σχηματίζονται, κατά κανόνα, μεταξύ κυττάρων του ίδιου τύπου (για παράδειγμα, μεταξύ κυττάρων του καρδιακού μυός).
Οι γενικές ιδιότητες των διεγερτικών ηλεκτρικών συνάψεων είναι:
Ταχύτητα (υπερβαίνει σημαντικά αυτή στις χημικές συνάψεις).
Η αδυναμία των ιχνών επιδράσεων κατά τη μεταφορά της διέγερσης (ως αποτέλεσμα αυτού, η άθροιση διαδοχικών σημάτων είναι πρακτικά αδύνατη σε αυτά).
Υψηλή αξιοπιστία μετάδοσης διέγερσης.
Οι διεγερτικές ηλεκτρικές συνάψεις μπορεί να προκύψουν υπό ευνοϊκές συνθήκες και να εξαφανιστούν κάτω από δυσμενείς. Για παράδειγμα, εάν ένα από τα κύτταρα που έρχονται σε επαφή είναι κατεστραμμένο, οι ηλεκτρικές συνάψεις του με άλλα κύτταρα εξαλείφονται. Αυτή η ιδιότητα ονομάζεται πλαστικότητα. Οι ηλεκτρικές συνάψεις μπορεί να είναι μονομερείς ή αμφίπλευρες.
Ηλεκτρική ανασταλτική σύναψη.Μαζί με τις ηλεκτρικές συνάψεις διεγερτικής δράσης, μπορεί να εμφανιστούν ηλεκτρικές ανασταλτικές συνάψεις. Το ανασταλτικό αποτέλεσμα συμβαίνει λόγω της δράσης του ρεύματος που προκαλείται από το δυναμικό δράσης της προσυναπτικής μεμβράνης. Το προσυναπτικό δυναμικό προκαλεί σημαντική υπερπόλωση του τμήματος και το υπερπολωτικό ρεύμα αναστέλλει αμέσως τη δημιουργία ενός δυναμικού δράσης στο αρχικό τμήμα του άξονα.
ΣΕ μικτές συνάψειςτο δυναμικό προσυναπτικής δράσης δημιουργεί ένα ρεύμα που εκπολώνει τη μετασυναπτική μεμβράνη μιας τυπικής χημικής σύναψης, όπου οι προ- και μετασυναπτικές μεμβράνες δεν προσκολλώνται σφιχτά μεταξύ τους. Έτσι, σε αυτές τις συνάψεις, η χημική μετάδοση χρησιμεύει ως απαραίτητος ενισχυτικός μηχανισμός.
Τροφικές επιρροές που μεταδίδονται μέσω των συνάψεων.Εκτός από τη μετάδοση διεγερτικών και ανασταλτικών σημάτων που είναι λειτουργικά σημαντικά, οι συνάψεις παρέχουν τροφικός (δηλαδή, επηρεάζουν την ανάπτυξη και τη διαφοροποίηση) αλληλεπιδράσεις των κυττάρων που έρχονται σε επαφή, που πραγματοποιούνται με τη βοήθεια τροφικών παραγόντων πρωτεϊνικής φύσης, πιθανώς επίσης συσσωρευμένων σε κυστίδια. Αυτοί οι παράγοντες παρέχουν μεταβολική διατήρηση της απαραίτητης δομής και ιδιοτήτων αυτών των κυττάρων. Διμερείς τροφικές αλληλεπιδράσεις έχουν προταθεί σε όλες τις συνάψεις, αλλά έχουν μελετηθεί κυρίως σε σκελετικές νευρομυϊκές συνάψεις σπονδυλωτών. Η μυϊκή απονεύρωση οδηγεί στην απώλεια των μυϊκών ινών της διαφοροποίησης που επιτυγχάνεται στην οντογένεση.
μετασυναπτικές διαδικασίες.Οι αλλαγές στο δυναμικό της μετασυναπτικής μεμβράνης ως αποτέλεσμα της ενεργοποίησης της σύναψης ονομάζονται συναπτικό δυναμικό. Διεγερτικό μετασυναπτικό δυναμικό (VPSP) εμφανίζεται σε εκπολωτικές συνάψεις και οφείλεται σε ταυτόχρονη αύξηση της διαπερατότητας της μεμβράνης για τα ιόντα Na + και K +. Τα ρεύματα που προκύπτουν σε αυτή την περίπτωση είναι αντίθετα κατευθυνόμενα (νάτριο - μέσα στο κύτταρο, κάλιο - έξω). Το MF μετατοπίζεται προς την αποπόλωση μέχρι μια τιμή ίση με το ήμισυ του αθροίσματος των δυναμικών ισορροπίας E Na και E K . Ανασταλτικά μετασυναπτικά δυναμικά (IPSP) είναι αλλαγές υπερπόλωσης στο μαγνητικό πεδίο (μέχρι -80-90 mV) και οφείλονται στο άνοιγμα διαύλων για ιόντα K+ (που εξέρχονται από το κύτταρο) ή ιόντα Cl- (που εισέρχονται στο κύτταρο) ή και για τα δύο ιόντα στο την ίδια ώρα. Το πλάτος των συναπτικών δυναμικών εξαρτάται από την ποσότητα του μεσολαβητή που απελευθερώνεται (ο αριθμός των κβαντών) και, επομένως, αυτές οι αντιδράσεις είναι βαθμιαίος σε αντίθεση με το ΠΔ. Αυτή η κωδικοποίηση πλάτους του σήματος συχνότητας πραγματοποιείται στον μετασυναπτικό νευρώνα, με εξαίρεση την αξονική περιοχή του, στην οποία υπάρχει επιστροφή στην κωδικοποίηση συχνότητας λόγω του AP που διαδίδεται κατά μήκος της ίνας του άξονα.
Η εκπόλωση που δημιουργείται από το EPSP μπορεί να υπερβεί σημαντικά, να φτάσει ή να παραμείνει κάτω από το όριο διέγερσης (σημειωμένες με τελείες). Το εύρος της υπερπόλωσης λόγω της ανάπτυξης του IPSP εξαρτάται από τη συχνότητα του προσυναπτικού AP και επηρεάζει τη συχνότητα της δραστηριότητας υποβάθρου του μετασυναπτικού κυττάρου.
Επειδή η κατεύθυνση των ιοντικών ρευμάτων εξαρτάται από τη βαθμίδα του ηλεκτροχημικού δυναμικού ενός δεδομένου ιόντος, τότε το πλάτος και η πολικότητα του συναπτικού σήματος αλλάζουν με μια αλλαγή στο μαγνητικό πεδίο. Η τιμή του, στην οποία αλλάζει το πρόσημο της συναπτικής δράσης, ονομάζεται δυνατότητα αντιστροφής. Η εξάπλωση των συναπτικών δυναμικών οφείλεται μόνο στις φυσικές ιδιότητες της κυτταρικής μεμβράνης και επομένως συμβαίνει με εξασθένηση (μείωση του πλάτους). Τα συναπτικά δυναμικά που προκύπτουν σε διαφορετικές συνάψεις μπορούν να αλληλεπιδράσουν μεταξύ τους, υπακούοντας στους κανόνες της αλγεβρικής άθροισης. Η αποπόλωση που προκαλείται από το άθροισμα των EPSP φέρνει το MP πιο κοντά στην τιμή κατωφλίου για διέγερση. Αντίθετα, η επιβολή του ΤΣΣΠ το απομακρύνει. Ο βαθμός αποπόλωσης ή υπερπόλωσης της μεμβράνης εξαρτάται από την «αντιπαράθεση» μεταξύ των ιοντικών αγωγιμοτήτων και των ρευμάτων που ενεργοποιούνται κατά την EPSP και την IPSP. Ο κύριος ρόλος στην ενσωμάτωση των σημάτων που έρχονται στο κύτταρο παίζεται από ειδικά τμήματα της νευρωνικής μεμβράνης: οι διακλαδιζόμενοι κόμβοι του δενδριτικού δέντρου, η σωματική μεμβράνη και ο λόφος του άξονα. Τις περισσότερες φορές, οι διεγερτικές συνάψεις εντοπίζονται στην δενδριτική μεμβράνη, ενώ οι ανασταλτικές συνάψεις εντοπίζονται, κατά κανόνα, στη σωματική μεμβράνη. Η τελική ολοκλήρωση όλων των εισόδων λαμβάνει χώρα στο συλλογικό άξονα του άξονα. Δεδομένου ότι οι μετασυναπτικές διεργασίες συμβαίνουν σε διαφορετικές αποστάσεις από τη ζώνη ενεργοποίησης και διαδίδονται παθητικά, η συμβολή τους στην ενοποιητική έξοδο θα εξαρτηθεί από τον εντοπισμό των συναπτικών επαφών.
Αυτή η αλληλεπίδραση επηρεάζεται από τις γεωμετρικές σχέσεις μεταξύ διεγερτικών και ανασταλτικών συνάψεων που βρίσκονται σε διαφορετικά μέρη των δενδριτών, καθώς και από τα χαρακτηριστικά της διάδοσης του ηλεκτροτονικού ρεύματος κατά μήκος αυτών των δενδριτών. Σε αυτή την περίπτωση, στον μετασυναπτικό νευρώνα, κατά τη δημιουργία του AP, μπορεί να εμφανιστούν τα αποτελέσματα της πρόσθεσης και της αφαίρεσης σημάτων εισόδου που έχουν τις ίδιες ή διαφορετικές (διεγερτικές και ανασταλτικές) μεθόδους.
Μετάδοση σήματος από μη παλμικούς νευρώνες.Ορισμένα νευρικά κύτταρα δεν δημιουργούν ΑΡ υπό φυσιολογικές συνθήκες (αυτή η ικανότητα, χαρακτηριστική για τους νευρώνες, εμφανίζεται μετά από ορισμένους τύπους χημικής έκθεσης). Παραδείγματα τέτοιων κυττάρων σε σπονδυλωτά είναι οι φωτοϋποδοχείς και τα κοκκώδη κύτταρα των οσφρητικών βολβών. Ωστόσο, αυτοί οι νευρώνες διατηρούν το πρότυπο: ο νευροδιαβιβαστής απελευθερώνεται μόνο κατά την εκπόλωση του προσυναπτικού κυττάρου.
Η διέγερση, έχοντας προκύψει σε ένα τμήμα της μεμβράνης ενός διεγέρσιμου κυττάρου, έχει την ικανότητα να εξαπλώνεται. Μια μακρά διαδικασία ενός νευρώνα - ένας άξονας (νευρική ίνα) εκτελεί στο σώμα μια συγκεκριμένη λειτουργία διέγερσης σε μεγάλες αποστάσεις.
Νόμοι διέγερσηςκατά μήκος των νευρικών ινών
Ο νόμος της ανατομικής και φυσιολογικής συνέχειας -Η διέγερση μπορεί να εξαπλωθεί κατά μήκος της νευρικής ίνας μόνο εάν είναι μορφολογικά και λειτουργικά ανέπαφη.
Ο νόμος της διμερούς διεξαγωγής της διέγερσης- η διέγερση που συμβαίνει σε ένα μέρος του νεύρου εξαπλώνεται και προς τις δύο κατευθύνσεις από τον τόπο προέλευσής του. Στο σώμα, η διέγερση διαδίδεται πάντα κατά μήκος του άξονα από το κυτταρικό σώμα (ορθόδρομα).
Ο Νόμος της Απομονωμένης Συμπεριφοράς- η διέγερση που διαδίδεται κατά μήκος της ίνας που είναι μέρος του νεύρου δεν μεταδίδεται σε γειτονικές νευρικές ίνες.
Μοτίβα του τοπικούκαι σκορπώντας ενθουσιασμό
Ηλεκτροτονικό δυναμικό (τοπική διέγερση)
• διαδίδεται κατά μήκος των νευρικών ινών με εξασθένηση (με μείωση), δηλ. το πλάτος της τοπικής απόκρισης πέφτει γρήγορα με την αύξηση της απόστασης από τον τόπο εμφάνισής της.
• λόγω της εξασθένησης, η τοπική απόκριση εκτείνεται σε μικρές αποστάσεις (όχι περισσότερες από 2 cm).
• Η τοπική διέγερση εξαπλώνεται παθητικά, χωρίς να ξοδεύεται η κυτταρική ενέργεια.
• ο μηχανισμός διάδοσης της τοπικής διέγερσης είναι παρόμοιος με τη διάδοση του ηλεκτρικού ρεύματος στους αγωγούς. Αυτός ο τύπος διάδοσης ονομάζεται ηλεκτροτονικό.
Δυνατότητα δράσης (διέγερση εξάπλωσης)
• διαδίδεται κατά μήκος των νευρικών ινών χωρίς εξασθένηση, το πλάτος του δυναμικού δράσης είναι το ίδιο σε οποιαδήποτε απόσταση από τον τόπο προέλευσής του.
• Η απόσταση στην οποία εκτείνεται το δυναμικό δράσης περιορίζεται μόνο από το μήκος της νευρικής ίνας.
• Η διάδοση του δυναμικού δράσης είναι μια ενεργή διαδικασία κατά την οποία αλλάζει η κατάσταση των καναλιών ιόντων ινών, απαιτείται ενέργεια ATP για την αποκατάσταση των διαμεμβρανικών ιοντικών βαθμίδων.
• ο μηχανισμός αγωγής του δυναμικού δράσης είναι πιο περίπλοκος από τον μηχανισμό διάδοσης της τοπικής διέγερσης.
Μυελινωμένες και μη μυελινωμένες νευρικές ίνες
ίνες μυελίνης. Ορισμένες νευρικές ίνες υφίστανται μυελίωση κατά την εμβρυογένεση: τα λεμοκύτταρα (κύτταρα Schwann) πρώτα αγγίζουν τον άξονα και μετά τον περιβάλλουν (Εικ. 1, Α, Β). Η μεμβράνη των λεμοκυττάρων τυλίγεται γύρω από τον άξονα σαν ρολό, σχηματίζοντας μια πολυστρωματική έλικα (θηκάρι μυελίνης) (Εικ. 1, C, D). Το περίβλημα μυελίνης δεν είναι συνεχές - σε όλο το μήκος της νευρικής ίνας σε ίση απόσταση το ένα από το άλλο, υπάρχουν μικρά σπασίματα σε αυτό (ο Ranvier παρεμποδίζει). Στην περιοχή των τομών, ο άξονας στερείται θήκης μυελίνης.
μη μυελινωμένες ίνες. Η μυελίνωση των άλλων ινών τελειώνει στα αρχικά στάδια της εμβρυϊκής ανάπτυξης. Ένας ή περισσότεροι άξονες βυθίζονται στο λεμοκύτταρο. τα περιβάλλει πλήρως ή εν μέρει, αλλά δεν σχηματίζει πολυστρωματικό περίβλημα μυελίνης (Εικ. 1ε).
Ο μηχανισμός αγωγής της διέγερσης κατά μήκος μη μυελινωμένων νευρικών ινών
Σε ηρεμία, ολόκληρη η εσωτερική επιφάνεια της μεμβράνης της νευρικής ίνας φέρει αρνητικό φορτίο και η εξωτερική πλευρά της μεμβράνης είναι θετική. Το ηλεκτρικό ρεύμα μεταξύ της εσωτερικής και της εξωτερικής πλευράς της μεμβράνης δεν ρέει, καθώς η λιπιδική μεμβράνη έχει υψηλή ηλεκτρική αντίσταση.
Κατά την ανάπτυξη του δυναμικού δράσης, εμφανίζεται μια αναστροφή φορτίου στην διεγερμένη περιοχή της μεμβράνης (Εικ. 2, Α). Ένα ηλεκτρικό ρεύμα αρχίζει να ρέει στα όρια των διεγερμένων και μη διεγερμένων τμημάτων (Εικ. 2, Β). Το ηλεκτρικό ρεύμα ερεθίζει το πλησιέστερο τμήμα της μεμβράνης και το φέρνει σε κατάσταση διέγερσης (Εικ. 2, C), ενώ τα προηγουμένως διεγερμένα τμήματα επιστρέφουν σε κατάσταση ηρεμίας (Εικ. 2, D). Έτσι, το κύμα διέγερσης καλύπτει όλα τα νέα τμήματα της μεμβράνης των νευρικών ινών.
Ο μηχανισμός αγωγής της διέγερσης κατά μήκος των νευρικών ινών της μυελίνης
Σε μια μυελινωμένη νευρική ίνα, τα τμήματα της μεμβράνης που καλύπτονται από το περίβλημα της μυελίνης είναι μη διεγέρσιμα. διέγερση μπορεί να συμβεί μόνο σε περιοχές της μεμβράνης που βρίσκονται στην περιοχή των τομέων του Ranvier.Με την ανάπτυξη του AP, εμφανίζεται μια αντιστροφή του φορτίου της μεμβράνης σε έναν από τους κόμβους του Ranvier (Εικ. 3, Α). Ένα ηλεκτρικό ρεύμα προκύπτει μεταξύ του ηλεκτραρνητικού και του ηλεκτροθετικού τμήματος της μεμβράνης, το οποίο ερεθίζει γειτονικά τμήματα της μεμβράνης (Εικ. 3, Β). Ωστόσο, μόνο ένα τμήμα της μεμβράνης στην περιοχή του επόμενου κόμβου του Ranvier μπορεί να εισέλθει στην κατάσταση διέγερσης (Εικ. 3γ). Έτσι, η διέγερση εξαπλώνεται σε όλη τη μεμβράνη με αλματώδη (αλατώδη) τρόπο από τη μια αναχαίτιση του Ranvier στην άλλη.
Ταξινόμηση νευρικών ινών
Οι νευρικές ίνες ποικίλλουν σε διάμετρο και βαθμό μυελίνωσης. Όσο μεγαλύτερη είναι η διάμετρος της νευρικής ίνας και ο βαθμός μυελίνωσης, τόσο μεγαλύτερος είναι ο ρυθμός αγωγής της διέγερσης. Οι ίνες με διαφορετικούς ρυθμούς αγωγιμότητας εκτελούν διαφορετικές φυσιολογικές λειτουργίες. Οι νευρικές ίνες χωρίζονται σε 6 τύπους, τα χαρακτηριστικά των οποίων δίνονται στον Πίνακα. 4.1.
Πίνακας 4.1. Τύποι νευρικών ινών, οι ιδιότητες και ο λειτουργικός τους σκοπός
Διάμετρος (μm) |
μυελίνωση |
Ταχύτητα αγωγιμότητας (m/s) |
Λειτουργικός σκοπός |
|
Ίνες κινητήρα του σωματικού NS; αισθητηριακές ίνες ιδιοϋποδοχέων |
||||
Αισθητηριακές ίνες των υποδοχέων του δέρματος |
||||
Αισθητήριες ίνες ιδιοϋποδοχέων |
||||
Αισθητηριακές ίνες θερμοϋποδοχέων, αλγοϋποδοχέων |
||||
Προγαγγλιακές ίνες του συμπαθητικού NS |
||||
λείπει |
Μεταγαγγλιακές ίνες του συμπαθητικού NS. αισθητήριες ίνες θερμοϋποδοχέων, αλγοϋποδοχείς, μερικοί μηχανοϋποδοχείς |
Οι νευρικές ίνες όλων των ομάδων έχουν κοινές ιδιότητες:
• Οι νευρικές ίνες είναι πρακτικά ακούραστες.
• Οι νευρικές ίνες έχουν υψηλή αστάθεια, δηλαδή μπορούν να αναπαράγουν το δυναμικό δράσης με πολύ υψηλή συχνότητα.

