20.01.2022
Fibre nervoase nemielinice și mielinice. Conceptul de „fibră nervoasă”
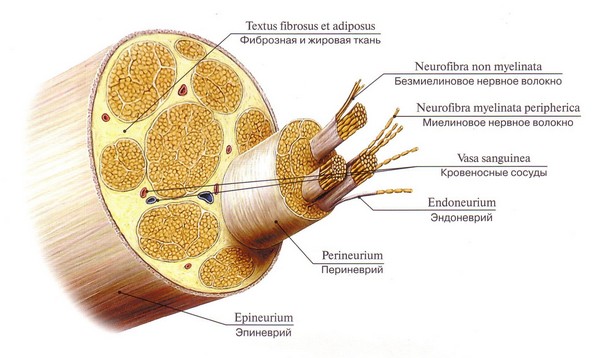
O fibră nervoasă este un proces alungit de neuroni acoperiți cu lemocite și o teacă de mielină sau non-mielinică. Funcția sa principală este conducerea impulsurilor nervoase. În sistemul nervos periferic și central predomină fibrele nervoase pulpoase (mielinice) care inervează mușchii scheletici; Fibrele care nu au înveliș se numesc cilindri axiali goi.
Fibra nervoasă se bazează pe procesul neuronului, care formează un fel de axă. În exterior, este înconjurat de o teacă de mielină cu o bază lipidică biomoleculară, constând dintr-un număr mare de spire de mesaxon, care se înfășoară spiralat în jurul axei neuronale. Astfel, are loc mielinizarea fibrelor nervoase.
Fibrele nervoase mielinice ale sistemului periferic sunt acoperite suplimentar de sus de celule Schwann auxiliare care susțin axonul și hrănesc corpul neuronului. Suprafața membranei pulpe are intervale - interceptări ale lui Ranvier, în aceste locuri cilindrul axial este atașat de membrana exterioară Schwann.
Stratul de mielină nu are proprietăți conductoare electric, ele au interceptări. Excitația are loc în intervalul Ranvier cel mai apropiat de locul expunerii la un stimul extern. Impulsul se transmite brusc, de la o interceptare la alta, aceasta asigura o viteza mare de propagare a impulsului.
Fibrele nervoase de mielină reglează metabolismul în țesutul muscular, au o rezistență ridicată la curentul bioelectric.
Lacunele lui Ranvier generează și amplifică impulsuri. Fibrele sistemului nervos central nu au membrană Schwann; această funcție este îndeplinită de oligodendroglia.
Țesuturile amielinice au mai mulți cilindri axiali, nu au strat de mielină și interceptări, sunt acoperite cu celule Schwann de sus, între ele și cilindri se formează spații sub formă de fante. Fibrele au o izolație slabă, permit propagarea unui impuls de la un proces al unui neuron la altul, contactul cu mediul în întregime, viteza de conducere a impulsului este mult mai mică decât cea a fibrelor pulpoase, în timp ce organismul are nevoie de mai multă energie.
Trunchiurile nervoase mari se formează din procesele pulpoase și necarnoase ale neuronilor, care, la rândul lor, se ramifică în mănunchiuri mai mici și se termină cu terminații nervoase (receptor, motor, sinapse).
Terminațiile nervoase sunt capătul fibrelor nervoase mielinizate și nemielinizate, care formează contacte interneuronale, receptorii și terminațiile motorii.
Principii de clasificare

Diferite tipuri de fibre nervoase au o rată inegală de conducere a impulsurilor excitatoare, aceasta depinde de diametrul lor, durata potențialului de acțiune și gradul de mielinizare. Există o relație direct proporțională între viteză și diametrul fibrei.
Metodă structural-funcțională de clasificare a fibrelor nervoase Erlanger-Gasser în funcție de viteza de conducere a impulsului nervos:
- Fibre nervoase mielinice din grupa A: α, β, Υ și δ. Cel mai mare diametru și cel mai gros înveliș sunt țesuturile α - 20 microni, au o viteză bună de conducere a pulsului - 120 m/s. Aceste țesuturi inervează sursa de excitație de la coloana măduvei spinării la receptorii mușchilor scheletici, tendoane și sunt responsabile pentru senzațiile tactile.
Tipurile rămase de fibre au un diametru mai mic (12 microni), viteza impulsului. Aceste țesuturi transmit semnale de la organele interne, surse de durere în SNC.
- Fibrele de mielină din grupa B aparțin. Viteza totală de conducere a impulsului este de 14 m/s, potențialul de acțiune este de 2 ori mai mare decât cel al fibrelor din grupa A. Teaca de mielină este slab exprimată.
- Fibrele nemielinizate din grupa C au un diametru foarte mic (0,5 microni) și o viteză de excitație (6 m/s). Aceste țesuturi inervează sistemul nervos simpatic. Acest grup include și fibre care conduc impulsurile din centrele durerii, frigului, căldurii și presiunii.
Procesele neuronilor sunt împărțite în aferente și eferente. Primul tip asigură transmiterea impulsurilor de la receptorii tisulari la sistemul nervos central. Al doilea tip transmite excitația de la sistemul nervos central către receptorii tisulari.
Clasificarea funcțională a fibrelor nervoase de tip aferent conform Lloyd-Hunt:
Demienilizare
Procesul de demielinizare a fibrelor nervoase este o deteriorare patologică a tecii de mielină, care provoacă disfuncția țesuturilor. Patologia este cauzată de procese inflamatorii, tulburări metabolice, neuroinfectie, intoxicație sau ischemie tisulară. Mielina este înlocuită cu plăci fibroase, ceea ce duce la afectarea conducerii impulsurilor.
Primul tip de demielinizare este mielinopatia cauzată de reacții autoimune ale organismului, boala Canavan, sindromul Guillain-Barré, amiotrofia Charcot-Marie-Tooth.
Al doilea tip este mielinoclastia. Patologia se caracterizează printr-o predispoziție ereditară la distrugerea tecii de mielină (boala Binswanger).
Boli demielinizante

Bolile care conduc la distrugerea tecii de mielină sunt cel mai adesea de natură autoimună, o altă cauză poate fi tratamentul cu antipsihotice sau o predispoziție ereditară. Distrugerea stratului lipidic determină o scădere a vitezei de conducere a impulsurilor de stimul.
Bolile sunt împărțite în cele care afectează sistemul nervos central și patologii care afectează rețeaua periferică. Boli care afectează activitatea sistemului nervos central:
- Mielopatia măduvei spinării apare ca urmare a comprimării fibrelor de mielină de către hernii intervertebrale, tumori, fragmente osoase, după. La pacienți, sensibilitatea și forța musculară în zona afectată scad, apare pareza brațelor sau picioarelor, activitatea intestinelor și a sistemului urinar este perturbată și se dezvoltă atrofia mușchilor extremităților inferioare.
- Leucodistrofia creierului provoacă leziuni ale substanței albe. Pacienții au o coordonare afectată a mișcărilor, nu pot menține echilibrul. Se dezvoltă slăbiciune musculară, apar convulsii involuntare, un tic nervos. Înrăutățirea treptată a memoriei, a abilităților intelectuale, a vederii și a auzului. În etapele ulterioare, apar orbire, surditate, paralizie completă și dificultăți la înghițirea alimentelor.
- Leucoencefalopatia cu focală mică a creierului afectează cel mai adesea bărbații cu vârsta peste 60 de ani. Principalele motive sunt hipertensiunea arterială și predispoziția ereditară. Pacienții au memorie și atenție afectate, există letargie, dificultăți de vorbire. Mersul încetinește, coordonarea mișcărilor este perturbată, apare incontinența urinară, pacientului îi este greu să înghită alimente.
- Sindromul de demielinizare osmotică se caracterizează prin descompunerea tecilor de mielină în țesuturile creierului. Pacienții au o tulburare a aparatului de vorbire, o senzație constantă de somnolență, depresie sau iritabilitate, mutism, pareză a tuturor membrelor. În stadiile incipiente ale bolii, procesul de demielinizare este reversibil.
- Scleroza multiplă se manifestă prin amorțeală a unuia sau două membre, pierderea parțială sau completă a vederii, durere la mișcarea ochilor, amețeli, oboseală, tremor la nivelul membrelor, tulburări de coordonare a mișcărilor, furnicături în diferite părți ale corpului.
- Boala Devic este o boală autoimună inflamatorie care afectează nervul optic și măduva spinării. Simptomele includ diferite grade de deficiență vizuală, până la orbire, parapareză, tetrapareză, funcționare afectată a organelor pelvine.
Simptomele bolilor depind de zona de deteriorare a fibrelor de mielină. Puteți identifica procesul de demielinizare folosind tomografie computerizată, terapie prin rezonanță magnetică. Semne de deteriorare a sistemului nervos periferic sunt găsite pe electromiografie.
Mielinizarea căilor vizuale aferenteîncepe în corpul geniculat lateral în luna a cincea de gestație și se termină prin naștere la placa cribriformă. Oligodendrocitele responsabile pentru mielinizarea fibrelor SNC sunt în mod normal absente în retină. Studiile histologice au confirmat prezența oligodendrocitelor și mielinei presupuse în zonele fibrelor nervoase mielinizate și absența acestora în afara acestor zone.
La autopsia fibrelor de mielina retinele sunt detectate la aproximativ 1% din ochi și la 0,3-0,6% dintre pacienții oftalmici în timpul unei examinări de rutină.
fibre nervoase mielinice de obicei apar ca fascicule striate longitudinal la polii superiori si inferiori ai discului. În același timp, datorită proeminenței părților modificate ale discului și suprapunerii marginilor discului și a vaselor de sânge subiacente prin fibre, anomalia poate simula edemul discului optic.
La distal se formează fibrele formă neregulată a evantaiului. În zona de mielinizare, sunt uneori vizibile mici fisuri sau zone ale fundului normal. În 17-20% din cazuri sunt detectate fibre nervoase mielinice bilaterale. În 19% din cazuri, acestea nu intră în contact cu capul nervului optic. Ocazional, fascicule izolate de fibre nervoase mielinice sunt detectate la periferia retinei pe partea nazală a ONH.
Patogeneza fibrelor de mielină rămâne neclar, dar animalele cu o lamină cribroasă slabă sau absentă prezintă de obicei excavare fiziologică profundă și mielinizare extinsă a fibrelor nervoase retiniene, în timp ce animalele cu o lamină cribriformă bine dezvoltată prezintă un disc optic complet plat (OND) și lipsă de mielinizare a retinei, care sugerează mai multe mecanisme posibile de patogeneză:
1. Prezența unui defect în placa cribriformă, care permite oligodendrocitelor să pătrundă în retină și să producă mielină.
2. Este posibil ca numărul de axoni să fie mic în comparație cu dimensiunea canalului scleral și să existe suficient spațiu pentru ca mielinizarea să continue în cavitatea oculară. La ochii cu fascicule de fibre nervoase mielinice izolate periferic, întreruperea timpului laminei cribrose permite oligodendrocitelor să intre în retină și să migreze în stratul de fibre nervoase până când ajung într-o zonă cu densitate relativ scăzută a fibrelor nervoase unde mielinizarea continuă.
3. Dezvoltarea târzie a plăcii cribriforme poate crea condiții pentru migrarea oligodendrocitelor în țesuturile oculare.
Mielinizarea extinsă unilaterală (sau, rar, bilaterală) a fibrelor nervoase poate fi însoțită de miopie mare și ambliopie refractivă. La acești pacienți, mielina acoperă cea mai mare parte, dacă nu toată, circumferința discului. În regiunea maculară (deși lipsită de mielină), sunt de obicei detectate și anomalii - o slăbire a reflexului sau dispersia pigmentului. Starea zonei maculare este probabil cel mai precis predictor al eficacității terapiei ocluzive.
fibre nervoase mielinice poate fi însoțită de sindromul Gorlin (nevi cu celule bazale multiple) și vitreoretinopatie autosomal dominantă, însoțită de tulburări vizuale congenitale, mielinizare extinsă bilaterală a stratului de fibre nervoase retiniene, degenerare severă a vitrosului, miopie ridicată, distrofie retiniană, orbire nocturnă și deformări ale membrelor.
fibre nervoase mielinice poate fi moștenit în mod autosomal dominant. Au fost descrise cazuri izolate de fibre nervoase mielinice asociate cu nervul optic lung anormal (oxicefalie), defecte ale laminei cribrose (disc optic oblic), disgeneza segmentului anterior și neurofibromatoză de tip 2. Deși fibrele nervoase mielinice pot coexista cu neurofibromatoza, mulți autori consideră această combinație discutabilă.
Ocazional zone mielinizarea fibrelor nervoase poate apărea după copilărie și chiar la vârsta adultă. În astfel de cazuri, traumatismul ochiului (traumatism contondent al ochiului într-un caz și fenestrarea tecilor nervului optic în altul) este probabil cauza obișnuită a acestei anomalii.
Este posibil ca astfel de pagube există un defect în placa cribriformă suficient pentru pătrunderea oligodendrocitelor în retină. Fibrele nervoase mielinice pot dispărea atunci când axonii sunt deteriorați.
Miopie unilaterală de grad înalt cu fibre nervoase mielinice. fibre nervoase mielinice.
fibre nervoase mielinice. A. Gradul de lumină. B. Grad sever.
Procesele neuronilor sunt aproape întotdeauna acoperite cu o teacă (mielină). Excepția o constituie terminațiile libere ale unor procese. Procesul împreună cu teaca se numește „fibră nervoasă”.
Fibra nervoasă este formată din: cilindru de osie- excrescere a unei celule nervoase: axon sau dendrita
Teaca gliala, înconjurând cilindrul axial sub forma unui cuplaj. În SNC, este format din oligodendroglia, iar în SNP de celule Schwann (neurolemocitele sunt un tip de oligodendroshlia).
Fibrele nervoase sunt clasificate în nemielinice și mielinizate (având o teacă de mielină).
Fibrele nervoase nemielinice fac parte din sistemul nervos autonom și sunt reprezentate de axonii neuronilor efectori. Sunt și în sistemul nervos central, dar în cantități mai mici.
Structura: În centru se află nucleul unui oligodendrocit (lemocit), iar de-a lungul periferiei 10-20 de cilindri axiali pătrund în citoplasma acestuia. Astfel de fibre nervoase sunt numite și „fibre de tip cablu”. Când cilindrul axial este scufundat în citoplasma oligodendrocitelor, secțiunile plasmolemei acestuia din urmă se apropie una de alta și se formează un mezenter - „mesaxon” sau o membrană dublă. De la suprafață, fibra nervoasă este acoperită cu o membrană bazală.
Fibrele nervoase mielinice fac parte din SNC, diviziunile somatice ale SNP și diviziunile preganglionare ale sistemului nervos autonom. Ele pot conține atât axoni, cât și dendrite ale neuronilor.
Structura: Cilindrul axial este întotdeauna 1, situat în centru. Cochilia are 2 straturi: interior (mielina) si exterior (neurolema), reprezentate de nucleul si citoplasma celulei Schwann. În exterior există o membrană de subsol. Stratul de mielină este format din mai multe straturi ale membranei oligodendrocitelor (lemocitelor). Membrana este răsucită concentric în jurul cilindrului axial. De fapt, acesta este un mesaxon foarte alungit. Mesaxonii formează procese citoplasmatice linguale.
Procesul de mielinizare este formarea tecii de mielină. Apare în stadiile târzii ale embriogenezei și în primele luni după naștere.
Este de remarcat faptul că există caracteristici ale mielinizării în SNC: 1 oligodendrocit formează o teacă de mielină în jurul mai multor cilindri axiali (cu ajutorul mai multor procese care se rotesc). Nu există membrană bazală.
Structura fibrei de mielină.
Mielina este întreruptă în mod regulat la nodurile lui Ranvier. Distanța dintre interceptări este de 0,3 - 1,5 nm. În zona de interceptare, se efectuează trofismul cilindrului axial. Mielina are crestături pe suprafața sa. Aceste secțiuni de tăiere a mielinei cresc flexibilitatea fibrei nervoase și reprezintă o „rezervă” pentru întindere. Nu există crestături în SNC.
Mielina este colorată cu coloranți pentru lipide: Sudan, acid osmic.
Funcțiile mielinei:
O creștere a vitezei de conducere a impulsului nervos. Fibrele fără mielină au o viteză de 1-2 m/s, în timp ce fibrele mielinice au o viteză de 5-120 m/s.
Canalele Na sunt concentrate în zona interceptărilor, unde apar curenți bioelectrici. Ei sar de la o interceptare la alta. Aceasta este conducerea saltativă, adică conducerea unui impuls în sărituri.
Mielina este un izolator care limitează intrarea curenților care se răspândesc în jur.
Diferența de structură a fibrelor mielinice și nemielinizate.
fibra nemielinizata fibra mielinizata
Mai mulți cilindri de osie 1 cilindru de osie
Cilindri axiali – axoni
Nucleul oligodendrocitelor se află în centru Nucleul și citoplasma oligodendrocitelor sunt la periferia fibrei
Mesaxonii sunt scurti Mesaxonii sunt răsuciți în mod repetat în jurul cilindrului axial, se formează teaca de mielină
Na- canale pe toată lungimea cilindrului axial Na- canale numai în nodurile lui Ranvier
Structura nervului periferic.
Nervul este format din fibre mielinice și nemielinice grupate în mănunchiuri. Conține atât fibre aferente, cât și eferente.
Mecanisme de conducere a impulsului nervos.
Sinapsele sunt conexiuni intercelulare speciale folosite pentru a transfera un semnal de la o celulă la alta.
Zonele de contact ale neuronilor sunt foarte apropiate unele de altele. Dar, cu toate acestea, între ele rămâne adesea un decalaj sinaptic care îi separă. Lățimea despicăturii sinaptice este de ordinul câtorva zeci de nanometri.
Pentru ca neutronii să funcționeze cu succes, este necesar să se asigure izolarea lor unul de celălalt, iar interacțiunea dintre ei este asigurată de sinapse.
Sinapsele acționează ca amplificatori ai semnalelor nervoase de-a lungul drumului lor. Efectul este obținut prin faptul că un impuls electric de putere relativ scăzută eliberează sute de mii de molecule de neurotransmițători, conținute anterior în multe vezicule sinaptice. O salvă de molecule mediatoare acționează sincron pe o zonă mică a neuronului controlat, unde sunt concentrați receptorii postsinaptici - proteine specializate care convertesc semnalul acum dintr-o formă chimică într-una electrică.
În prezent, principalele etape ale procesului de eliberare a mediatorului sunt bine cunoscute. Un impuls nervos, adică un semnal electric, apare într-un neuron, se propagă de-a lungul proceselor sale și ajunge la terminațiile nervoase. Transformarea sa într-o formă chimică începe cu deschiderea canalelor ionice de calciu în membrana presinaptică, a cărei stare este controlată de câmpul electric al membranei. Acum ionii de calciu preiau rolul de purtători de semnal. Ei intră prin canalele deschise din interiorul terminației nervoase. O concentrație puternic crescută de ioni de calciu în apropierea membranei pentru o perioadă scurtă de timp activează mașina moleculară pentru eliberarea mediatorului: veziculele sinaptice merg în locurile fuziunii lor ulterioare cu membrana exterioară și, în cele din urmă, își aruncă conținutul în spațiul despicăturii sinaptice. .
Transmiterea sinaptică se realizează printr-o secvență de două procese separate spațial: presinaptic pe o parte a golului sinaptic și postsinaptic pe cealaltă (Fig. 3). Terminațiile proceselor neuronului de control, supunând semnalelor electrice care au ajuns la ele, eliberează o substanță intermediară specială (mediator) în spațiul despicăturii sinaptice. Moleculele mediatoare difuzează destul de repede prin fanta sinaptică și excită un semnal electric de răspuns în celula controlată (un alt neuron, fibre musculare, unele celule ale organelor interne). Aproximativ o duzină de substanțe diferite cu greutate moleculară mică acționează ca mediator:
acetilcolină (un ester al aminoalcoolului colinei și acidului acetic); glutamat (anion al acidului glutamic); GABA (acid gamma-aminobutiric); serotonină (un derivat al aminoacidului triptofan); adenozină etc.
Ele sunt sintetizate preliminar de neuronul presinaptic din materii prime disponibile și relativ ieftine și depozitate până când sunt utilizate în vezicule sinaptice, unde, ca în recipiente, sunt închise porțiuni identice ale mediatorului (câteva mii de molecule într-o veziculă).
diagrama sinapselor
Deasupra - o secțiune a terminației nervoase, limitată de membrana presinaptică, în care sunt încorporați receptorii presinaptici; veziculele sinaptice din interiorul terminației nervoase sunt umplute cu un mediator și sunt în diferite grade de pregătire pentru eliberarea acestuia; membranele veziculare și membrana presinaptică conțin proteine presinaptice. Mai jos - o secțiune a unei celule controlate, în membrana postsinaptică a cărei receptori postsinaptici sunt încorporați.
Sinapsele sunt un obiect convenabil pentru reglarea fluxurilor de informații. Nivelul de amplificare a semnalului în timpul transmiterii acestuia prin sinapsă poate fi ușor crescut sau micșorat prin modificarea cantității de mediator eliberat, până la interzicerea completă a transmiterii informațiilor. Teoretic, acest lucru se poate realiza prin țintirea oricăreia dintre etapele de eliberare a mediatorului.
Fibre de mielină. O parte din fibrele nervoase în timpul embriogenezei suferă mielinizare: lemocitele (celulele Schwann) ating mai întâi axonul și apoi îl învăluie. Membrana lemocitelor se înfășoară în jurul axonului ca o rolă, formând o spirală multistratificată (teaca de mielină). Teaca de mielină nu este continuă - de-a lungul întregii lungimi a fibrei nervoase la o distanță egală una de cealaltă, există mici întreruperi în ea (interceptări Ranvier). În zona interceptărilor, axonul este lipsit de înveliș de mielină.
fibre nemielinice. Mielinizarea altor fibre se termină în stadiile incipiente ale dezvoltării embrionare. Unul sau mai mulți axoni sunt scufundați în lemocit; le înconjoară complet sau parțial, dar nu formează o teacă de mielină multistratificată.
Mecanismul de conducere a excitației de-a lungul fibrelor nervoase nemielinice
În repaus, întreaga suprafață interioară a membranei fibrei nervoase poartă o sarcină negativă, iar partea exterioară a membranei este pozitivă. Curentul electric între partea interioară și exterioară a membranei nu curge, deoarece. membrana lipidica are o rezistenta electrica mare.In timpul dezvoltarii potentialului de actiune in regiunea excitata a membranei are loc o reversie a sarcinii. La limita zonei excitate și neexcitate, începe să curgă un curent electric. Un curent electric irită cea mai apropiată secțiune a membranei și o aduce într-o stare de excitare, în timp ce secțiunile excitate anterior revin la o stare de repaus. Astfel, unda de excitație acoperă toate secțiunile noi ale membranei fibrei nervoase.
Mecanismul de conducere a excitației de-a lungul fibrelor nervoase de mielină
Într-o fibră nervoasă mielinică, porțiunile membranei acoperite de teaca de mielină sunt neexcitabile; excitația poate apărea numai în zonele membranei situate în zona nodurilor lui Ranvier Odată cu dezvoltarea AP într-unul dintre nodurile lui Ranvier, sarcina membranei este inversată. Între secțiunile electronegativă și electropozitivă ale membranei ia naștere un curent electric, care irită secțiunile învecinate ale membranei. Cu toate acestea, doar o secțiune a membranei din zona următorului nod al lui Ranvier poate intra în starea de excitație. Astfel, excitația se răspândește pe membrană într-o manieră sărită (saltatorie) de la o interceptare a lui Ranvier la alta.
FUNCȚIILE UNUI NEURON
Materialul pentru construirea sistemului nervos central și a conductorilor acestuia este țesutul nervos, care constă din două componente - celule nervoase (neuroni) și neuroglia. Principalele elemente funcționale ale SNC sunt neuronii: în corpul animalelor există aproximativ 50 de miliarde dintre ei, dintre care doar o mică parte este localizată în părțile periferice ale corpului.
Neuronii reprezintă 10 - 15% din numărul total de elemente celulare din sistemul nervos. Cea mai mare parte a acestuia este ocupată de celule neurogliale. La animalele superioare, în procesul ontogenezei postnatale, neuronii diferențiați nu se divid. (cum se credea anterior). Neuronii diferă semnificativ în formă (piramidală, rotundă, stelata, ovale), mărime (de la 5 la 150 microni), număr de procese, dar au și proprietăți comune.
În unele părți ale creierului există neuroni care produc granule de secreție de natură mucoproteică sau glicoproteică. Au atât caracteristici fiziologice ale neuronilor, cât și ale celulelor glandulare. Aceste celule sunt numite neurosecretoare. Funcția neuronilor este de a percepe semnale de la receptori sau de la alte celule nervoase, de a stoca și procesa informații și de a transmite impulsuri nervoase către alte celule - nervoase, musculare sau secretoare. În consecință, există o specializare a neuronilor. Aceștia sunt împărțiți în 3 grupe: neuroni senzitivi (senzoriali, aferenți), neuroni asociativi (intermediari, intercalari), neuroni motori (efectori).
Neuroglia. Cea mai mare parte a țesutului nervos este alcătuită din elemente gliale care îndeplinesc funcții auxiliare și umplu aproape întreg spațiul dintre neuroni. Din punct de vedere anatomic, celulele neurogliale din creier (oligodendrocite și astrocite) și celulele Schwann din sistemul nervos periferic se disting printre ele. Oligodendrocitele și celulele Schwann formează teci de mielină în jurul axonilor. Între celulele gliale și neuroni există goluri de 15 - 20 nm lățime, care comunică între ele, formând un spațiu interstițial umplut cu lichid. Prin acest spațiu are loc un schimb de substanțe între neuron și celulele gliale, precum și furnizarea de oxigen și substanțe nutritive neuronilor prin difuzie. Celulele gliale, aparent, îndeplinesc doar funcții de susținere și de protecție în SNC și nu sunt, așa cum era de așteptat, sursa lor de nutriție sau deținătorii de informații.
Celulele gliale diferă de neuroni prin proprietățile membranei: reacţionează pasiv la curentul electric, membranele lor nu generează un impuls de propagare. Există joncțiuni strânse (zone cu rezistență scăzută) între celulele neurogliale care asigură o conexiune electrică directă. Potențialul de membrană al celulelor gliale este mai mare decât cel al neuronilor și depinde în principal de concentrația ionilor K+ din mediu.
Când concentrația de K + crește în timpul activității active a neuronilor în spațiul extracelular, o parte din aceasta este absorbită de elementele gliale depolarizate. Această funcție de tamponare a gliei asigură o concentrație extracelulară relativ constantă de K+. Celulele gliale - astrocite - sunt situate între corpurile neuronilor și peretele capilarelor, procesele lor sunt în contact cu peretele acestora din urmă. Aceste procese perivasculare sunt elemente ale barierei hemato-encefalice. Celulele microgliale îndeplinesc o funcție fagocitară, numărul lor crește brusc atunci când țesutul cerebral este deteriorat.
Fiziologia sinapselor.
Sinapsele chimice au o despicatură sinaptică relativ largă (20 - 50 nm) și rezistență mare a membranelor sinaptice. Terminalul nervos presinaptic conține un număr mare de vezicule - vezicule sinaptice - circa 50 nm în diametru, umplut cu mediator.
Mecanismul transmiterii excitației în sinapsele excitatorii chimice. În sinapsele cu transmisie chimică, excitația este transmisă folosind mediatori(intermediari). Mediat ori - acestea sunt substanțe chimice care asigură transmiterea excitației în sinapse. Mediatorii, în funcție de natura lor, sunt împărțiți în mai multe grupuri:
monoamine(acetilcolina, dopamina, norepinefrina, serotonina etc.);
Aminoacizi(acid gamma-aminobutiric - GABA, acid glutamic, glicină etc.);
neuropeptide(substanța P, endorfine, neurotensină, ACTH, angiotensină, vasopresină, somatostatina etc.). Mediatorul în formă moleculară este situat în veziculele îngroșării presinaptice (placă sinaptică), unde intră:
Din regiunea perinucleară a neuronului folosind transport axonal rapid (axocurent);
Datorită sintezei unui mediator care apare în terminalele sinaptice din produsele sale de clivaj;
Datorită recaptării neurotransmițătorului din fanta sinaptică într-o formă neschimbată.
Varietate de sisteme mediatoare. Principiul Dale: un neuron, de regulă, sintetizează și folosește un mediator în toate terminalele sale. Este posibil ca un neuron să folosească mai mulți mediatori (comedianți) , dar aparent în aceeași combinație (acest principiu este în prezent contestat, vezi mai jos pentru sistemele de mediator) . Ca o consecință a principiului Dale, pot fi luate în considerare următoarele prevederi:
1) Semnul acțiunii sinaptice este determinat nu de mediator, ci de proprietățile receptorilor de pe celula postsinaptică.
2) Receptorii de pe celulele care sunt postsinaptice la un singur neuron presinaptic pot diferi din punct de vedere farmacologic și pot controla diferite canale ionice.
3) O celulă postsinaptică poate avea mai mult de un tip de receptor pentru un mediator dat și fiecare dintre acești receptori poate controla un mecanism de conducere ionică diferit.
Datorită acestor trei proprietăți, celulele pot exercita acțiuni sinaptice opuse atât asupra celulelor postsinaptice diferite, cât și asupra aceleiași.
Mediatorii identificați până în prezent la animale și la oameni constituie un grup destul de eterogen de substanțe. Monoamine: acetilcolina, dopamina, norepinefrina, serotonina (5-hidroxitriptamina, 5-HT), histamina. Aminoacizi: acid gama-aminobutiric (GABA), acid glutamic, glicină, taurină, etc. Neuropeptide: methenkefalin, leuenkefalin, endorfină, oxitocină ,vasopresină si etc . Sinapsele corespunzătoare sunt numite, de exemplu, colinergic, serotoninergic, noradrenergic etc. Un număr mare de mediatori corespunde unui număr mare de receptori postsinaptici: receptorii colinergici , adrenoreceptori, receptori GABA etc. În fiecare grup de receptori, există o varietate de subtipuri ale acestora, de exemplu, receptorii de acetilcolină nicotinici și muscarinici (H-XR și, respectiv, M-XR).
Divizarea receptorilor se realizează pe baza diferențelor în proprietățile lor farmacologice: diferite agonişti (substanţe care imită efectul unui mediator) şi antagonişti (substanţe care împiedică manifestarea efectului mediatorului). De exemplu, pentru N-ChR, agonistul este nicotina , antagonişti - tubocurarina (izolat din veninul de curare), bungarotoxină (izolat din veninul unui şarpe din genul Bungarus). Există trei tipuri de receptori de glutamat bazați pe agonişti: quisqualat (tip AMPA), kainat și NMDA (N-metil-D-aspartat) -tip. Unii receptori mediatori (în special, adrenoreceptori și receptori pentru multe neuropeptide) nu sunt asociați cu canalele ionice (receptori ionotropi) , dar cu o enzimă membranară (receptori metabotropi) , De exemplu, adenilat ciclază . Acesta din urmă, odată activat de un mediator, catalizează transformarea multor molecule adenozin trifosfat (ATP) ciclic adenozin monofosfat (cAMP) - mecanism de amplificare. CAMP, fiind un al doilea mesager, activează multe enzime din celulă, în special protein kinaze si astfel stimuleaza metabolismul celular. AMPc este distrus fosfodiesteraza. Sistemul de adenilat ciclază este similar cu sistemul guanilat ciclază, sistemul fosfolipază C etc. (Fig. 14). Formarea cGMP din forma neciclică este catalizată de guanilat ciclază , a cărui activitate este stimulată de oxidul nitric. Moleculele acestuia din urmă se formează în timpul dezaminării argininei și formării citrulinei sub acțiunea oxid nitric sintaza. Activitatea acestei enzime este, la rândul ei, reglată de complex Ca 2+ -calmodulină. Astfel, de exemplu, receptorii de glutamat, care declanșează fluxul de intrare al ionilor de Ca 2+, controlează fluctuațiile concentrației de cGMP în citoplasma neuronilor. În creierul vertebratelor, activitatea oxidului nitric sintazei se găsește în cerebel, cvadrigemina, striat și bulbul olfactiv.
Mecanismul sinapselor chimice:în timpul depolarizării terminalului presinaptic (cauzat de PD sau artificial), ionii de Ca 2+ intră în el din mediu, care stimulează procesul exocitoza - golirea veziculelor în fanta sinaptică.
Proteinele sinaptice și veziculare caracteristice sunt prezentate împreună cu receptorii și funcțiile lor presupusi. Sunt postulate secțiuni separate ale membranei veziculare pentru ancorarea veziculelor pe citoschelet), atașarea membranei veziculare de membrana presinaptică și eliberarea mediatorului prin porul rezultat. Mecanismele moleculare de atașare a veziculelor la membrana presinaptică și formarea porilor sunt probabil diferite. Unele dintre proteine sunt ținte ale neurotoxinelor care modifică eliberarea neurotransmițătorului. De exemplu, structura proteinelor sinaptobrevine veziculare (VAMP) este perturbată de toxinele tetanos și botulinice; veninul de păianjen latrotoxina se leagă de proteinele membranei presinaptice neurexinele și îmbunătățește golirea veziculelor.
1. Sinapsinele sunt proteine asociate veziculelor care se presupune că leagă veziculele sinaptice de citoscheletul terminației nervoase. 2. Atașarea, formarea porilor și golirea veziculelor sunt mediate de interacțiunile (indicate prin săgeți) ale diferitelor proteine veziculare și membranare. De exemplu, proteinele veziculare (sinaptotagmină și sinaptobrevine) și proteinele membranei plasmatice a terminației nervoase (sintaxine și neurexine) sunt implicate în formarea complexului de atașare. 3. Ce proteine - plasmatice sau veziculare formează porul de fuziune încă nu este clar. Probabil, acestea sunt sinaptofizina (care au alte funcții) și proteina membranei plasmatice, fizofilina. 4. Proteinele Rab pot fi implicate în transportul veziculelor în celulă și în atașarea acestora la membrana celulelor nervoase. Transportoarele veziculare joacă un rol în acumularea de neurotransmițători în veziculele sinaptice.
Simultan, Ca 2+ începe să fie îndepărtat din citoplasmă în mai multe moduri: legarea de proteine, absorbția mitocondrială și transportul activ. Ieșirea mediatorului depinde de depolarizarea terminalului și este de aproximativ 100 - 200 de vezicule, fiecare dintre ele conținând o porțiune (cuantică), corespunzătoare aproximativ 10 4 molecule. Moleculele mediatoare difuzează către membrana postsinaptică, unde interacționează cu receptorii membranei postsinaptice care reglează starea canalelor ionice. Această reglare poate fi directă (ca, de exemplu, în joncțiunea neuromusculară a mușchilor scheletici ai vertebratelor) sau poate include activarea sistemelor. secundar mediatori intracelulari (proteine G, cAMP).
În primul caz, sunt transmise semnale rapide de pornire, în al doilea, efecte mai lente pe termen lung. Direcția de schimbare a potențialului membranei postsinaptice (depolarizare sau hiperpolarizare) depinde în principal de deschiderea cărora canalele controlează receptorii postsinaptici. Unele dintre moleculele mediatoare pot interacționa cu receptori presinaptici, ceea ce duce la o modificare a MP a terminalului nervos și, în consecință, a cantității de mediator eliberat (feedback). Despicatură sinaptică este curățată de mediator în diferite moduri: dezactivare, hidroliză, recaptare în terminația presinaptică, difuzie, captare de către celulele gliale. Parte principală întârziere sinaptică - timpul de la sosirea unui impuls nervos până la dezvoltarea unui răspuns postsinaptic (0,2-0,5 ms) cade asupra procesului de secreție a neurotransmițătorului. Sinapsa chimică asigură transmiterea semnalului doar de la neuronul presinaptic la cel postsinaptic.
Cu stimulare ritmică frecventă în sinapsele chimice, o creștere a (relief) iar apoi atenuare (depresie) transmisie, adică urcă și apoi scade în amplitudinea potențialelor postsinaptice. Aceste fenomene sunt determinate în principal de modificări ale legăturii presinaptice. Au o dezvoltare deosebită în unele sinapse ale SNC, unde acţionează ca factori de plasticitate sinaptică. Prezența diferitelor tipuri de receptori în membrana postsinaptică poate determina dezvoltarea acestor fenomene separat, cum ar fi potențarea pe termen lung și depresia pe termen lung.
Sinapsele inhibitoare chimice. Aceste sinapse, în ceea ce privește mecanismul de transmitere a excitației, sunt similare cu sinapsele cu acțiune excitatoare. În sinapsele inhibitoare, un mediator (de exemplu, glicina) interacționează cu receptorii de pe membrana subsinaptică și deschide canalele de clorură în ea, ceea ce duce la mișcarea ionilor de clorură de-a lungul gradientului de concentrație în celulă și dezvoltarea hiperpolarizării pe membrana subsinaptică. . Există un așa-zis potenţial postsinaptic inhibitor(TPSP).
Anterior, se credea că fiecare mediator corespunde unei reacții specifice a celulei postsinaptice - excitare sau inhibare într-o formă sau alta. S-a stabilit acum că un mediator corespunde cel mai adesea nu unuia, ci mai multor receptori diferiți. De exemplu, acetilcolina din sinapsele neuromusculare ale mușchilor scheletici acționează asupra receptorilor H-colinergici (sensibili la nicotină), care deschid canale largi pentru sodiu (și potasiu), care generează EPSP (PKP) În sinapsele vago-cardiace, aceeași acetilcolină. acţionează asupra receptorilor M- colinergici (sensibili la muscarină), care deschid canale selective pentru ionii de potasiu, astfel încât aici se generează potenţial postsinaptic inhibitor (IPSP). În consecință, natura excitatoare sau inhibitorie a acțiunii mediatorului este determinată de proprietățile membranei subsinaptice (mai precis, de tipul de receptor), și nu de mediatorul în sine.
Proprietățile fiziologice ale sinapselor chimice. Sinapsele cu transmitere chimică a excitației au o serie de proprietăți comune:
Excitația prin sinapse se realizează doar într-o singură direcție (unilateral). Acest lucru se datorează structurii sinapsei: mediatorul este eliberat numai din îngroșarea presinaptică și interacționează cu receptorii membranei subsinaptice;
Transmiterea excitatiei prin sinapse este mai lenta decat prin fibra nervoasa - intarziere sinaptica;
Transferul excitației se realizează cu ajutorul unor mediatori chimici speciali - mediatori;
În sinapse, ritmul de excitație este transformat;
Sinapsele au labilitate scăzută;
Sinapsele sunt foarte obosite;
Sinapsele sunt foarte sensibile la substanțele chimice (inclusiv farmacologice).
Sinapsele electrice excitatorii. Pe lângă sinapsele cu transmisie chimică a excitației, sinapsele cu transmisie electrică se găsesc predominant în sistemul nervos central (SNC). Sinapsele electrice excitatorii se caracterizează printr-o despicatură sinaptică foarte îngustă și o rezistivitate foarte scăzută a membranelor pre- și postsinaptice adiacente, ceea ce asigură trecerea eficientă a curenților electrici locali. Rezistența scăzută, de regulă, este asociată cu prezența canalelor transversale care traversează ambele membrane, adică merg de la celulă la celulă (contact cu gol). Canalele sunt formate din molecule proteice (semimolecule) ale fiecăreia dintre membranele în contact, care sunt conectate complementar. Această structură este ușor de circulat pentru curent electric.
Diagrama transmiterii excitației într-o sinapsă electrică: curentul cauzat de potențialul de acțiune presinaptic irită membrana postsinaptică, unde apar EPSP și potențialul de acțiune. Canalele transversale unesc celulele nu numai electric, ci și chimic, deoarece sunt acceptabile pentru mulți compuși cu greutate moleculară mică. Prin urmare, sinapsele electrice excitatorii cu canale transversale se formează, de regulă, între celulele de același tip (de exemplu, între celulele mușchiului cardiac).
Proprietățile generale ale sinapselor electrice excitatorii sunt:
Viteza (depășește semnificativ pe cea din sinapsele chimice);
Slăbiciunea efectelor urmelor în timpul transferului de excitație (ca urmare a acestui fapt, însumarea semnalelor succesive este practic imposibilă în ele);
Fiabilitate ridicată a transmisiei excitației.
Sinapsele electrice excitatoare pot apărea în condiții favorabile și pot dispărea în condiții nefavorabile. De exemplu, dacă una dintre celulele în contact este deteriorată, sinapsele sale electrice cu alte celule sunt eliminate. Această proprietate se numește plasticitate. Sinapsele electrice pot fi unilaterale sau bilaterale.
Sinapsa inhibitorie electrică. Alături de sinapsele electrice de acțiune excitatoare pot apărea și sinapsele electrice inhibitorii. Efectul inhibitor apare datorita actiunii curentului cauzat de potentialul de actiune al membranei presinaptice. Potențialul presinaptic determină o hiperpolarizare semnificativă a segmentului iar curentul hiperpolarizant inhibă instantaneu generarea unui potențial de acțiune în segmentul inițial al axonului.
LA sinapse mixte potențialul de acțiune presinaptică creează un curent care depolarizează membrana postsinaptică a unei sinapse chimice tipice, unde membranele pre- și postsinaptice nu aderă strâns între ele. Astfel, în aceste sinapse, transmisia chimică servește ca un mecanism de întărire necesar.
Influențe trofice transmise prin sinapse. Pe lângă transmiterea semnalelor excitatorii și inhibitorii care sunt importante din punct de vedere funcțional, sinapsele oferă trofic (adică afectarea creșterii și diferențierii) interacțiuni ale celulelor aflate în contact, realizate cu ajutorul unor factori trofici de natură proteică, probabil acumulați și în vezicule. Acești factori asigură menținerea metabolică a structurii și proprietăților necesare acestor celule. Interacțiuni trofice bilaterale au fost sugerate în toate sinapsele, dar au fost studiate în principal în sinapsele neuromusculare ale scheletului de vertebrate. Denervarea musculară duce la pierderea fibrelor musculare a diferențierii realizate în ontogeneză.
procesele postsinaptice. Se numesc modificări ale potențialului membranei postsinaptice ca urmare a activării sinapselor potenţial sinaptic. Potenţial postsinaptic excitator (VPSP) apare în sinapsele depolarizante și se datorează unei creșteri simultane a permeabilității membranei pentru ionii Na + și K +. Curenții care apar în acest caz sunt direcționați opus (sodiu - în interiorul celulei, potasiu - afară). MF se deplasează spre depolarizare până la o valoare egală cu jumătate din suma potențialelor de echilibru E Na și E K . Potențiale postsinaptice inhibitoare (IPSP) sunt modificări de hiperpolarizare în câmpul magnetic (până la -80-90 mV) și se datorează deschiderii canalelor pentru ionii K + (care părăsesc celula), sau ionii Cl - (intra în celulă), sau pentru ambii ioni la acelasi timp. Amplitudinea potențialelor sinaptice depinde de cantitatea de mediator eliberat (numărul de cuante) și, astfel, aceste reacții sunt treptat spre deosebire de PD. Această codificare în amplitudine a semnalului de frecvență este efectuată în neuronul postsinaptic, cu excepția regiunii sale axonale, în care există o revenire la codificarea frecvenței datorită propagării AP de-a lungul fibrei axonale.
Depolarizarea creată de EPSP poate depăși semnificativ, atinge sau rămâne sub pragul de excitație (marcat cu puncte). Amplitudinea hiperpolarizării datorată dezvoltării IPSP depinde de frecvența AP presinaptică și afectează frecvența activității de fond a celulei postsinaptice.
pentru că direcția curenților ionici depinde de gradientul potențialului electrochimic al unui ion dat, apoi amplitudinea și polaritatea semnalului sinaptic se modifică odată cu modificarea câmpului magnetic. Se numește valoarea sa, la care semnul acțiunii sinaptice se schimbă potenţial de inversare. Răspândirea potențialelor sinaptice se datorează numai proprietăților fizice ale membranei celulare și, prin urmare, are loc cu atenuare (scăderea amplitudinii). Potențialele sinaptice care apar în diferite sinapse pot interacționa între ele, respectând regulile însumării algebrice. Depolarizarea cauzată de însumarea EPSP-urilor aduce MP mai aproape de valoarea de prag pentru excitație. Dimpotriva, impunerea TSSP il distanteaza. Gradul de depolarizare sau hiperpolarizare a membranei depinde de „confruntarea” dintre conductivitățile ionice și curenții activați în timpul EPSP și IPSP. Rolul principal în integrarea semnalelor care vin în celulă este jucat de secțiuni speciale ale membranei neuronale: nodurile de ramificare a arborelui dendritic, membrana somatică și dealul axonului. Cel mai adesea, sinapsele excitatorii sunt localizate pe membrana dendritică, în timp ce sinapsele inhibitoare sunt localizate, de regulă, pe membrana somatică. Integrarea finală a tuturor intrărilor are loc la coliculul axon. Deoarece procesele postsinaptice apar la distanțe diferite de zona de declanșare și se propagă pasiv, contribuția lor la ieșirea integrativă va depinde de localizarea contactelor sinaptice.
Această interacțiune este influențată de relațiile geometrice dintre sinapsele excitatorii și inhibitorii localizate în diferite părți ale dendritelor, precum și de caracteristicile propagării curentului electrotonic de-a lungul acestor dendrite. În acest caz, în neuronul postsinaptic, în timpul generării AP, pot apărea efectele adunării și scăderii semnalelor de intrare având aceleași sau diferite modalități (excitatorii și inhibitorii).
Transmiterea semnalului prin neuroni nepulsatori. Unele celule nervoase nu generează AP în condiții fiziologice (această abilitate, tipică neuronilor, se manifestă după anumite tipuri de expunere chimică). Exemple de astfel de celule la vertebrate sunt fotoreceptorii și celulele granulare ale bulbilor olfactiv. Cu toate acestea, acești neuroni păstrează modelul: neurotransmițătorul este eliberat numai la depolarizarea celulei presinaptice.
Excitația, care a apărut într-o secțiune a membranei unei celule excitabile, are capacitatea de a se răspândi. Un proces lung al unui neuron - un axon (fibră nervoasă) îndeplinește în organism o funcție specifică de conducere a excitației pe distanțe lungi.
Legile excitatieide-a lungul fibrelor nervoase
Legea continuitatii anatomice si fiziologice - excitația se poate răspândi de-a lungul fibrei nervoase numai dacă este intactă din punct de vedere morfologic și funcțional.
Legea conducerii bilaterale a excitației- excitația care are loc într-o parte a nervului se răspândește în ambele direcții de la locul de origine. În organism, excitația se propagă întotdeauna de-a lungul axonului din corpul celular (ortodrom).
Legea conduitei izolate- excitația care se propagă de-a lungul fibrei care face parte din nervul nu este transmisă fibrelor nervoase învecinate.
Modele ale localuluiși răspândind entuziasm
Potențial electrotonic (excitație locală)
• se propagă de-a lungul fibrelor nervoase cu atenuare (cu scăderea), adică amplitudinea răspunsului local scade rapid odată cu creșterea distanței de la locul apariției acestuia;
• din cauza atenuarii, raspunsul local se extinde pe distante scurte (nu mai mult de 2 cm);
• excitația locală se răspândește pasiv, fără a cheltui energia celulară;
• mecanismul de propagare a excitaţiei locale este similar cu propagarea curentului electric în conductori; Acest tip de propagare se numește electrotonice.
Potential de actiune (excitatie raspandita)
• se propaga de-a lungul fibrelor nervoase fara atenuare, amplitudinea potentialului de actiune este aceeasi la orice distanta de locul de origine;
• distanta pana la care se extinde potentialul de actiune este limitata doar de lungimea fibrei nervoase;
• propagarea potențialului de acțiune este un proces activ în timpul căruia starea canalelor ionice ale fibrei se modifică, este necesară energia ATP pentru restabilirea gradienților ionici transmembranari;
• mecanismul conducţiei potenţialului de acţiune este mai complex decât mecanismul de propagare a excitaţiei locale.
Fibre nervoase mielinice și nemielinice
fibre de mielina. Unele fibre nervoase suferă mielinizare în timpul embriogenezei: lemocitele (celulele Schwann) ating mai întâi axonul și apoi îl învăluie (Fig. 1, A, B). Membrana lemocitelor se înfășoară în jurul axonului ca o rolă, formând o spirală multistrat (teaca de mielină) (Fig. 1, C, D). Teaca de mielină nu este continuă - de-a lungul întregii lungimi a fibrei nervoase la o distanță egală una de cealaltă, există mici întreruperi în ea (interceptări Ranvier). În zona interceptărilor, axonul este lipsit de înveliș de mielină.
fibre nemielinice. Mielinizarea altor fibre se termină în stadiile incipiente ale dezvoltării embrionare. Unul sau mai mulți axoni sunt scufundați în lemocit; le înconjoară complet sau parțial, dar nu formează o teacă de mielină multistrat (Fig. 1e).
Mecanismul de conducere a excitației de-a lungul fibrelor nervoase nemielinice
În repaus, întreaga suprafață interioară a membranei fibrei nervoase poartă o sarcină negativă, iar partea exterioară a membranei este pozitivă. Curentul electric între partea interioară și exterioară a membranei nu circulă, deoarece membrana lipidică are o rezistență electrică ridicată.
În timpul dezvoltării potențialului de acțiune, are loc o reversie a sarcinii în regiunea excitată a membranei (Fig. 2, A). Un curent electric începe să curgă la limita secțiunilor excitate și neexcitate (Fig. 2, B). Curentul electric irită cea mai apropiată secțiune a membranei și o aduce într-o stare de excitație (Fig. 2, C), în timp ce secțiunile excitate anterior revin la o stare de repaus (Fig. 2, D). Astfel, unda de excitație acoperă toate secțiunile noi ale membranei fibrei nervoase.
Mecanismul de conducere a excitației de-a lungul fibrelor nervoase de mielină
Într-o fibră nervoasă mielinică, porțiunile membranei acoperite de teaca de mielină sunt neexcitabile; excitația poate apărea numai în zonele membranei situate în regiunea interceptărilor lui Ranvier.Odată cu dezvoltarea AP, are loc o inversare a sarcinii membranei într-unul dintre nodurile lui Ranvier (Fig. 3, A). Între secțiunile electronegativă și electropozitivă ale membranei ia naștere un curent electric, care irită secțiunile învecinate ale membranei (Fig. 3, B). Cu toate acestea, doar o porțiune a membranei din regiunea următorului nod al lui Ranvier poate intra în starea de excitație (Fig. 3c). Astfel, excitația se răspândește pe membrană într-o manieră sărită (saltatorie) de la o interceptare a lui Ranvier la alta.
Clasificarea fibrelor nervoase
Fibrele nervoase variază ca diametru și grad de mielinizare. Cu cât este mai mare diametrul fibrei nervoase și gradul de mielinizare, cu atât este mai mare viteza de conducere a excitației. Fibrele cu viteze de conducere diferite îndeplinesc diferite funcții fiziologice. Fibrele nervoase sunt împărțite în 6 tipuri, ale căror caracteristici sunt prezentate în tabel. 4.1.
Tabelul 4.1. Tipuri de fibre nervoase, proprietățile lor și scopul funcțional
Diametru (µm) |
mielinizare |
Viteza de conducere (m/s) |
Scop functional |
|
Fibrele motorii ale NS somatic; fibrele senzoriale ale proprioreceptorilor |
||||
Fibrele senzoriale ale receptorilor pielii |
||||
Fibrele senzoriale ale proprioreceptorilor |
||||
Fibre senzoriale ale termoreceptorilor, nociceptorilor |
||||
Fibrele preganglionare ale NS simpatic |
||||
este absent |
Fibre postganglionare ale NS simpatic; fibrele senzoriale ale termoreceptorilor, nociceptorilor, unor mecanoreceptori |
Fibrele nervoase din toate grupele au proprietăți comune:
• fibrele nervoase sunt practic neobosite;
• fibrele nervoase au o labilitate mare, adică pot reproduce potenţialul de acţiune cu o frecvenţă foarte mare.

