20.01.2022
Безмиелиновые и миелиновые нервные волокна. Понятие «нервное волокно»
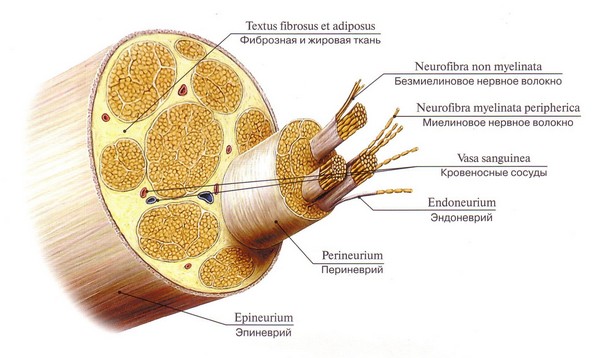
Нервное волокно – это удлиненный отросток нейронов, покрытый леммоцитами и миелиновой или безмиелиновой оболочкой. Основной его функцией является проводимость нервных импульсов. В периферической и центральной нервной системе преобладают мякотные (миелиновые) нервные волокна, которые иннервируют скелетную мускулатуру, безмякотные находятся в симпатическом отделе вегетативной системы и распространяются на внутренние органы. Волокна, не имеющие оболочки, называются голыми осевыми цилиндрами.
Нервное волокно имеет в основе отросток нейрона, который образует своеобразную ось. Снаружи он окружен миелиновой оболочкой с биомолекулярной липидной основой, состоящей из большого количества витков мезаксона, который по спирали накручивается на нейроновую ось. Таким образом, происходит миелинизация нервных волокон.
Миелиновые нервные волокна периферической системы сверху дополнительно покрыты вспомогательными Шванновскими клетками, поддерживающими аксон и питающими тело нейрона. Поверхность мякотной мембраны имеет интервалы – перехваты Ранвье, в этих местах осевой цилиндр прикрепляется к наружной Шванновской мембране.
Миелиновый слой не обладает электропроводящими свойствами, их имеют перехваты. Возбуждение происходит в ближайшем к месту воздействия внешнего раздражителя интервале Ранвье. Импульс передается скачкообразно, от одного перехвата к другому, это обеспечивает высокую скорость распространения импульса.
Миелиновые нервные волокна регулируют обмен веществ в мышечной ткани, обладают высоким сопротивлением по отношению к биоэлектрическому току.
Промежутки Ранвье генерируют и усиливают импульсы. У волокон центральной нервной системы нет Шванновской мембраны, эту функцию выполняют олигодендроглии.
Безмякотные ткани имеют несколько осевых цилиндров, у них нет миелинового слоя и перехватов, сверху покрыты Шванновскими клетками, между ними и цилиндрами образуются щелевидные пространства. Волокна имеют слабую изоляцию, допускают распространение импульса из одного отростка нейрона в другой, на всем протяжении контактируют с окружающей средой, скорость проведения импульсов гораздо ниже, чем у мякотных волокон, при этом организму требуется большее количество энергии.
Из мякотных и безмякотных отростков нейронов формируются крупные нервные стволы, которые, в свою очередь, разветвляются на более мелкие пучки и заканчиваются нервными окончаниями (рецепторные, двигательные, синапсы).
Нервные окончания – это конец миелиновых и безмиелиновых нервных волокон, который формирует межнейронные контакты, рецепторные и двигательные окончания.
Принципы классификации

Разные типы нервных волокон имеют неодинаковую скорость проведения импульсов возбуждения, это зависит от их диаметра, длительности потенциала действия и степени миелинизации. Существует прямо пропорциональная зависимость между скоростью и диаметром волокна.
Структурно-функциональный метод классификации нервных волокон Эрлангера-Гассера по скорости проведения нервных импульсов:
- Миелиновое нервное волокно группы А: α, β, Υи δ. Самый большой диаметр и толстую оболочку имеют ткани α – 20 мк, они обладают хорошей скорость проводимости импульсов – 120 м/сек. Эти ткани иннервируют источник возбуждения из столба спинного мозга к скелетным рецепторам мышц, сухожильям, отвечают за тактильные ощущения.
Остальные типы волокон имеют меньший диаметр (12 мк), скорость проведения импульса. Эти ткани передают сигналы от внутренних органов, источников боли в ЦНС.
- Миелиновые волокна группы В относятся к . Общая скорость проведения импульса составляет 14 м/сек, потенциал действия в 2 раза больше, чем у волокон группы А. Миелиновая оболочка слабо выражена.
- Безмиелиновые волокна группы С имеют очень маленький диаметр (0,5 мк) и скорость возбуждения (6 м/сек). Эти ткани иннервируют симпатическую нервную систему. К данной группе также относятся волокна, которые проводят импульсы от центров боли, холода, тепла и давления.
Отростки нейронов делят на афферентные и эфферентные. Первый тип обеспечивает передачу импульсов от рецепторов тканей в центральную нервную систему. Второй тип передает возбуждение от ЦНС к рецепторам тканей.
Функциональная классификация нервных волокон афферентного типа по Ллойду-Ханту:
Демиенилизация
Процесс демиелинизации нервных волокон – это патологическое повреждение миелиновой оболочки, которое вызывает нарушение функционирования тканей. Вызывают патологию воспалительные процессы, метаболические нарушения, нейроинфекция, интоксикация или ишемия тканей. Миелин замещается фиброзными бляшками, в результате нарушается проведение импульсов.
Первый тип демиелинизации – это миелинопатия, вызванная аутоиммунными реакциями организма, болезнью Канавана, синдромом Гийена-Барре, амиотрофией Шарко-Мари-Тута.
Второй тип – это миелинокластия. Патология характеризуется наследственной предрасположенностью к разрушению миелиновой оболочки (болезнь Бинсвангера).
Демиелинизирующие заболевания

Заболевания, приводящие к разрушению миелиновой оболочки, чаще всего имеют аутоиммунную природу, другой причиной может быть лечение нейролептиками или наследственная предрасположенность. Разрушение липидного слоя вызывает снижение скорости проведения импульсов раздражения.
Заболевания разделяют на те, которые затрагивают центральную нервную систему и патологии, повреждающие периферическую сеть. Болезни, которые влияют на работу ЦНС:
- Миелопатия спинного мозга возникает в результате сдавливания миелиновых волокон межпозвоночными грыжами, опухолями, костными осколками, после . У больных снижается чувствительность, мышечная сила в области поражения, возникают парезы рук или ног, нарушается работа кишечника, мочевыводящей системы, развивается атрофия мышц нижних конечностей.
- Лейкодистрофия головного мозга вызывает поражение белого вещества. У пациентов нарушена координация движений, они не могут держать равновесие. Развивается мышечная слабость, появляются непроизвольные судороги, нервный тик. Постепенно ухудшается память, интеллектуальные способности, зрение и слух. На поздних стадиях возникает слепота, глухота, полный паралич, трудности во время проглатывания пищи.
- Мелкоочаговая лейкоэнцефалопатия головного мозга чаще всего поражает мужчин старше 60 лет. Основными причинами является артериальная гипертензия и наследственная предрасположенность. У пациентов ухудшается память и внимание, появляется заторможенность, трудности с речью. Замедляется походка, нарушается координация движений, появляется недержание мочи, больному тяжело глотать пищу.
- Синдром осмотической демиелинизации характеризуется распадом миелиновых оболочек в тканях головного мозга. У больных отмечается расстройство речевого аппарата, постоянное чувство сонливости, депрессии или повышенная возбудимость, мутизм, парез всех конечностей. На ранних стадиях заболевания процесс демиелинизации обратим.
- Рассеянный склероз проявляется онемением одной или двух конечностей, частичная или полная потеря зрения, боль при движении глаз, головокружение, быстрая утомляемость, тремор конечностей, нарушение координации движений, покалывание в различных частях тела.
- Болезнь Девика – это воспалительный аутоиммунный недуг, который поражает зрительный нерв и ствол спинного мозга. К симптомам относится различная степень нарушения зрения, вплоть до слепоты, парапарезы, тетрапарезы, нарушение функционирования органов малого таза.
Симптомы заболеваний зависят от области поражения миелиновых волокон. Выявить процесс демиелинизации можно с помощью компьютерной томографии, магниторезонансной терапии. Признаки поражения периферической нервной системы обнаруживаются на электромиографии.
Миелинизация афферентных зрительных путей начинается в латеральном коленчатом теле на пятом месяце гестации и завершается к родам у решетчатой пластинки. Олигодендроциты, ответственные за миелинизацию волокон ЦНС, в норме в сетчатке отсутствуют. При гистологических исследованиях было подтверждено наличие предполагаемых олигодендроцитов и миелина в зонах миелиновых нервных волокон и их отсутствие вне этих зон.
При вскрытии миелиновые волокна сетчатки выявляются приблизительно в 1% глаз и у 0,3-0,6% офтальмологических пациентов при плановом обследовании.
Миелиновые нервные волокна обычно выглядят как продольно исчерченные пучки у верхнего и нижнего полюса диска . При этом из-за проминенции измененных частей диска и перекрывания волокнами краев диска и нижележащих кровеносных сосудов аномалия может симулировать отек ДЗН.
Дистально волокна формируют неправильную веерообразную фигуру . В пределах зоны миелинизации иногда видны мелкие щели или участки нормального глазного дна. В 17-20% случаев выявляются двусторонние миелиновые нервные волокна. В 19% случаев они не соприкасаются с диском зрительного нерва. Изредка на периферии сетчатки с носовой стороны от ДЗН выявляются изолированные пучки миелиновых нервных волокон.
Патогенез миелиновых волокон
остается не выясненным, но у животных со слабо выраженной или отсутствующей решетчатой пластинкой обычно имеется глубокая физиологическая экскавация и обширная миелинизация нервных волокон сетчатки, тогда как у животных с хорошо развитой решетчатой пластинкой наблюдается абсолютно плоский диск зрительного нерва (ДЗН) и отсутствие миелинизации сетчатки, что позволяет предположить несколько возможных механизмов патогенеза:
1. Наличие дефекта решетчатой пластинки, позволяющего олигодендроцитам проникать на сетчатку и вырабатывать миелин.
2. Возможно, количество аксонов по сравнению с размером склерального канала невелико, и имеется достаточно пространства, чтобы миелинизация продолжалась и в полости глаза. В глазах с периферическими изолированными пучками миелиновых нервных волокон, нарушение сроков формирования решетчатой пластинки позволяет олигодендроцитам проникать в сетчатку и мигрировать в слое нервных волокон, пока они не достигнут зоны относительно низкой плотности нервных волокон, где продолжается миелинизация.
3. Позднее развитие решетчатой пластинки может создавать условия для миграции олигодендроцитов в ткани глаза.
Обширная односторонняя (или, редко, двусторонняя) миелинизация нервных волокон может сопровождаться близорукостью высокой степени и рефракционной амблиопией. У таких пациентов миелин перекрывает большую часть, если не полностью, окружности диска. В макулярной области (хотя и свободной от миелина) также обычно выявляются аномалии - ослабление рефлекса или дисперсия пигмента. Состояние макулярной области, вероятно, является наиболее точным прогностическим фактором эффективности окклюзионной терапии.
Миелиновые нервные волокна могут сопутствовать синдрому Gorlin (множественных базальноклеточных невусов) и аутосомно-доминантной витреоретинопатии, сопровождающихся врожденным ухудшением зрения, двусторонней обширной миелинизацией слоя нервных волокон сетчатки, тяжелой дегенерацией стекловидного тела, близорукостью высокой степени, дистрофией сетчатки, ночной слепотой и деформациями конечностей.
Миелиновые нервные волокна могут наследоваться по аутосомно-доминантному типу. Описаны изолированные случаи миелиновых нервных волокон в сочетании с аномальной длинной зрительного нерва (при оксицефалии), дефектами решетчатой пластинки (косой диск зрительного нерва), дисгенезом переднего сегмента и нейрофиброматозом 2 типа. Хотя миелиновые нервные волокна могут сопутствовать нейрофиброматозу, многие авторы считают такое сочетание сомнительным.
Изредка зоны миелинизации нервных волокон могут возникать по прошествии младенческого возраста и даже во взрослом возрасте. Вероятно, в таких случаях обычной причиной этой аномалии является травма глаза (тупая травма глаза в одном случае и фенестрация оболочек зрительного нерва - в другом).
Возможно, при таких повреждениях возникает дефект решетчатой пластинки , достаточный для проникновения олигодендроцитов в сетчатку. Миелиновые нервные волокна могут исчезать при поражении аксонов.
Односторонняя близорукость высокой степени с миелиновыми нервными волокнами. Миелиновые нервные волокна.
Миелиновые нервные волокна.
А. Легкая степень. Б. Тяжелая степень.
Отростки нейронов почти всегда покрыты оболочкой (миелином). Исключение составляют свободные окончания некоторых отростков. Отросток вместе с оболочкой называется «нервное волокно».
Нервное волокно состоит из: Осевого цилиндра
– отросток нервной клетки: аксон или дендрит
Глиальной оболочки
, окружающей осевой цилиндр в виде муфты. В ЦНС она образована олигодендроглией, а в ПНС – шванновскими клетками (нейролеммоциты – разновидность олигодендрошлии).
Нервные волокна классифицируются на Безмиелиновые и Миелиновые (имеющие миелиновую оболочку).
Безмиелиновые нервные волокна являются частью вегетативной нервной системы и представлены аксонами эффекторных нейронов. Они есть и в ЦНС, но в меньшем количестве.
Строение: В центре находится ядро олигодендроцита (леммоцита), а по периферии в его цитоплазму проникают10-20 осевых цилиндров. Такие нервные волокна ещё называют «волокна кабельного типа». При погружении осевого цилиндра в цитоплазму олигодендроцита участки плазмолеммы последнего сближаются, и формируется брыжейка - «мезаксон» или сдвоенная мембрана. С поверхности нервное волокно покрыто базальной мембраной.
Миелиновые нервные волокна являются частью ЦНС, соматических отделов ПНС, а также преганглионарных отделов вегетативной нервной системы. Они могут содержать как аксоны, так и дендриты нейронов.
Строение: Осевой цилиндр всегда 1, расположен в центре. Оболочка имеет 2 слоя: внутренний (миелиноый) и наружный (нейролемма), представленный ядром и цитоплазмой шванновской клетки. Снаружи имеется базальная мембрана. Миелиновый слой представляет собой насколько слоёв мембраны олигодендроцита (леммоцита). Мембрана концентрически закручена вокруг осевого цилиндра. Фактически это очень удлинённый мезаксон. Мезаксоны образуют языковидные цитоплазматические отростки.
Процесс миелинизации – это образование миелиновой оболочки. Он происходит на поздних стадиях эмбриогенеза и в первые месяцы после рождения.
Стоит отметить, что в ЦНС есть особенности миелинизации: 1 олигодендроцит образует миелиновую оболочку вокруг нескольких осевых цилиндров (с помощью нескольких отростков, которые вращаются). Нет базальной мембраны.
Строение миелинового волокна.
Миелин регулярно прерывается в области узловых перехватов Ранвье. Расстояние между перехватами 0,3 – 1,5 нм. В области перехвата осуществляется трофика осевого цилиндра. Миелин имеет на своей поверхности насечки. Эти участки рассечения миелина увеличивают гибкость нервного волокна и являются «запасом» при растяжении. В ЦНС насечек нет.
Миелин окрашивается красителями на липиды: Судан, Осмиевая кислота.
Функции миелина:
Увеличение скорости проведения нервного импульса. У безмиелиновых волокон скорость 1-2 м/с, а у миелиновых – 5-120 м/с.
В области перехватов сосредоточены Na-каналы, где возникают биоэлектрические токи. Они перескакивают от 1 перехвата к другому. Это - сальтаторное проведение, то есть проведение импульса скачками.
Миелин – изолятор, который ограничивает вхождение токов, распространяющихся вокруг.
Различие в строении миелинового и безмиелинового волокна.
Безмиелиновое волокно Миелиновое волокно
Несколько осевых цилиндров 1 осевой цилиндр
Осевые цилиндры - аксоны Осевыми цилиндрами могут быть те, и др. Осевые цилиндры толще, чем в безмиелиновых волокнах
Ядро олигодендроцита – в центре Ядро и цитоплазма олигодендроцита – на периферии волокна
Мезаксоны короткие Мезаксон многократно закручен вокруг осевого циландра, образуется миелиновая оболочка
Na- каналы по всей длине осевого цилиндра Na- каналы только в перехватах Ранвье
Строение периферического нерва.
Нерв состоит из миелиновых и безмиелиновых волокон, сгруппированных в пучки. Он содержит и афферентные, и эфферентные волокна.
Механизмы проведения нервного импульса.
Синапсы - это специальные межклеточные соединения, используемые для перехода сигнала из одной клетки в другую.
Контактирующие участки нейронов очень тесно прилегают друг к другу. Но все же между ними зачастую остается разделяющая их синаптическая щель. Ширина синаптической щели составляет порядка нескольких десятков нанометров.
Чтобы нейтроны успешно функционировали, необходимо обеспечить их обособленность друг от друга, а взаимодействие между ними обеспечивают синапсы.
Синапсы выполняют функцию усилителей нервных сигналов на пути их следования. Эффект достигается тем, что один относительно маломощный электрический импульс освобождает сотни тысяч молекул медиатора, заключенных до того во многих синаптических пузырьках. Залп молекул медиатора синхронно действует на небольшой участок управляемого нейрона, где сосредоточены постсинаптические рецепторы - специализированные белки, которые преобразуют сигнал теперь уже из химической формы в электрическую.
В настоящее время хорошо известны основные этапы процесса освобождения медиатора. Нервный импульс, т. е. электрический сигнал, возникает в нейроне, распространяется по его отросткам и достигает нервных окончаний. Его преобразование в химическую форму начинается с открывания в пресинаптической мембране кальциевых ионных каналов, состояние которых управляется электрическим полем мембраны. Теперь роль носителей сигнала берут на себя ионы кальция. Они входят через открывшиеся каналы внутрь нервного окончания. Резко возросшая на короткое время примембранная концентрация ионов кальция активизирует молекулярную машину освобождения медиатора: синаптические пузырьки направляются к местам их последующего слияния с наружной мембраной и, наконец, выбрасывают свое содержимое в пространство синаптической щели.
Синаптическая передача осуществляется последовательностью двух пространственно разобщенных процессов: пресинаптического по одну сторону синаптической щели и постсинаптического по другую (рис. 3). Окончания отростков управляющего нейрона, повинуясь пришедшим в них электрическим сигналам, высвобождают в пространство синаптической щели специальное вещество-посредник (медиатор). Молекулы медиатора достаточно быстро диффундируют через синаптическую щель и возбуждают в управляемой клетке (другом нейроне, мышечном волокне, некоторых клетках внутренних органов) ответный электрический сигнал. В роли медиатора выступает около десятка различных низкомолекулярных веществ:
ацетилхолин (эфир аминоспирта холина и уксусной кислоты);глутамат (анион глутаминовой кислоты);ГАМК (гамма-аминомасляная кислота);серотонин (производное аминокислоты триптофана);аденозин и др.
Они предварительно синтезируются пресинаптическим нейроном из доступного и относительно дешевого сырья и хранятся вплоть до использования в синаптических пузырьках, где, словно в контейнерах, заключены одинаковые порции медиатора (по несколько тысяч молекул в одном пузырьке).
Схема синапса
Вверху - участок нервного окончания, ограниченный пресинаптической мембраной, в которую встроены пресинаптические рецепторы; синаптические пузырьки внутри нервного окончания наполнены медиатором и находятся в разной степени готовности к его освобождению; мембраны пузырьков и пресинаптическая мембрана содержат пресинаптические белки. Внизу - участок управляемой клетки, в постсинаптическую мембрану которой встроены постсинаптические рецепторы
Синапсы - удобный объект регулирования потоков информации. Уровень усиления сигнала при его передаче через синапс можно легко увеличить или уменьшить, изменяя количество освобождаемого медиатора, вплоть до полного запрета на передачу информации. Теоретически это можно осуществить путем направленного воздействия на любой из этапов высвобождения медиатора.
Миелиновые волокна . Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты (шванновские клетки) сначала прикасаются к аксону, а затем окутывают его. Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки.
Механизм проведения возбуждения по безмиелиновым нервным волокнам
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, т.к. липидная мембрана имеет высокое электрическое сопротивление.Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда. На границе возбужденного и невозбужденного участка начинает протекать электрический ток. Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения, в то время как ранее возбужденные участки возвращаются в состояние покоя. Таким образом, волна возбуждение охватывает все новые участки мембраны нервного волокна.
Механизм проведения возбуждения по миелиновым нервным волокнам
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только участках мембраны, расположенных в области перехватов Ранвье.При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны. Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны. Однако, в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье. Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
ФУНКЦИИ НЕЙРОНА
Материалом для построения ЦНС и ее проводников является нервная ткань, состоящая из двух компонентов - нервных клеток (нейронов) и нейроглии. Основными функциональными элементами ЦНС являются нейроны: в теле животных их содержится примерно 50 млрд, из которых лишь небольшая часть расположена на периферических участках тела.
Нейроны составляют 10 - 15 % общего числа клеточных элементов в нервной системе. Основную же часть ее занимают клетки нейроглии. У высших животных в процессе постнатального онтогенеза дифференцированные нейроны не делятся (так считалось ранее) . Нейроны существенно различаются по форме (пирамидные, круглые, звездчатые, овальные), размерами (от 5 до 150 мкм), количеству отростков, однако они имеют и общие свойства.
В некоторых отделах мозга имеются нейроны, которые вырабатывают гранулы секрета мукопротеидной или гликопротеидной природы. Они обладают одновременно физиологическими признаками нейронов и железистых клеток. Эти клетки называются нейросекреторными. Функция нейронов заключается в восприятии сигналов от рецепторов или других нервных клеток, хранении и переработке информации и передаче нервных импульсов к другим клеткам - нервным, мышечным или секреторным. Соответственно имеет место специализация нейронов. Их подразделяют на 3 группы: чувствительные (сенсорные, афферентные) нейроны, ассоциативные (промежуточные,вставочные) нейроны, двигательные (эффекторные) нейроны.
Нейроглия. Основную массу нервной ткани составляют глиальные элементы, выполняющие вспомогательные функции и заполняющие почти все пространство между нейронами. Анатомически среди них различают клетки нейроглии в мозге (олигодендроциты и астроциты) и шванновские клетки в периферической нервной системе. Олигодендроциты и шванновские клетки формируют вокруг аксонов миэлиновые обалочки. Между глиальными клетками и нейронами имеются щели шириной 15 - 20 нм, которые сообщаются друг с другом, образуя интерстициальное пространство, заполненное жидкостью. Через это пространство происходит обмен веществ между нейроном и глиальными клетками, а также снабжение нейронов кислородом и питательными веществами путем диффузии. Глиальные клетки, по-видимому, выполняют лишь опорные и защитные функции в ЦНС, а не являются, как предполагалось, источником их питания или хранителями информации.
По свойствам мембраны глиальные клетки отличаются от нейронов: они пассивно реагируют на электрический ток, их мембраны не генерируют распространяющегося импульса. Между клетками нейроглии существуют плотные контакты (участки низкого сопротивления), которые обеспечивают прямую электрическую связь. Мембранный потенциал глиальных клеток выше, чем у нейронов, и зависит главным образом от концентрации ионов К+ в среде.
Когда при активной деятельности нейронов во внеклеточном пространстве увеличивается концентрация К+, часть его поглощается деполяризованными глиальными элементами. Эта буферная функция глии обеспечивает относительно постоянную внеклеточную концентрацию К+. Клетки глии - астроциты - расположены между телами нейронов и стенкой капилляров, их отростки контактируют со стенкой последних. Эти периваскулярные отростки являются элементами гематоэнцефалического барьера. Клетки микроглии выполняют фагоцитарную функцию, число их резко возрастает при повреждении ткани мозга.
Физиология синапсов.
Химические синапсы имеют относительно широкую синаптическую щель (20 - 50 нм) и высокое сопротивление синаптических мембран. В пресинаптической нервной терминали находится большое число пузырьков - синаптических везикул - диаметром около 50 нм, заполненных медиатором.
Механизм передачи возбуждения в химических возбуждающих синапсах . В синапсах с химической передачей возбуждение передается с помощью медиаторов (посредников). Медиаторы - это химические вещества, которые обеспечивают передачу возбуждения в синапсах. Медиаторы в зависимости от их природы делятся на несколько групп:
моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.);
Аминокислоты (гамма-аминомасляная кислота - ГАМК, глутаминовая кислота, глицин и др.);
нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.). Медиатор в молекулярном виде находится в пузырьках пресинаптического утолщения (синаптической бляшке), куда он поступает:
Из околоядерной области нейрона с помощью быстрого аксонального транспорта (аксотока);
За счет синтеза медиатора, протекающего в синаптических терминалях из продуктов его расщепления;
За счет обратного захвата медиатора из синаптической щели в неизменном виде.
Многообразие медиаторных систем. Принцип Дейла: один нейрон, как правило, синтезирует и использует один медиатор во всех своих терминалях. Возможно использование нейроном нескольких медиаторов (комедиаторы) , но, по-видимому, в одном и том же сочетании (в настоящее время этот принцип оспаривается, о медиаторных системах см. ниже) . Как следствия принципа Дейла можно рассматривать следующие положения:
1) Знак синаптического действия определяется не медиатором, а свойствами рецепторов на постсинаптической клетке.
2) Рецепторы на клетках, являющихся постсинаптическими по отношению к одному пресинаптическому нейрону, могут фармакологически различаться и могут контролировать разные ионные каналы.
3) Одна постсинаптическая клетка может иметь более одного типа рецепторов для данного медиатора, и каждый из этих рецепторов может контролировать отличный от других механизм ионной проводимости.
Вследствие этих трех свойств клетки могут оказывать противоположные синаптические действия как на различные постсинаптические клетки, так и на одну и ту же.
Медиаторы, выявленные к настоящему времени у животных и человека, составляют довольно разнородную группу веществ. Моноамины: ацетилхолин, дофамин, норадреналин, серотонин (5-гидрокситриптамин, 5-ГТ), гистамин. Аминокислоты: гамма-аминомасляная кислота (ГАМК), глутаминовая кислота, глицин, таурин и др. Нейропептиды: метэнкефалин, лейэнкефалин, эндорфин, окситоцин ,вазопрессин и др. Соответствующие синапсы называют, например,холинэргические, серотонинэргические, норадренэргические и т.д. Большому количеству медиаторов соответствует большое количество постсинаптических рецепторов: холинорецетпоры , адренорецепторы, ГАМК-рецепторы и т.д. В пределах каждой группы рецепторов существует разнообразие их подтипов, например никотиновые и мускариновые холинорецепторы (Н-ХР и М-ХР, соответственно).
Разделение рецепторов проводится на основе различий их фармакологических свойств: разные агонисты (вещества, имитирующие эффект медиатора) и антагонисты (вещества, препятствующие проявлению эффекта медиатора). Например, для Н-ХР агонист - никотин, антагонисты - тубокурарин (выделен из яда кураре), бунгаротоксин (выделен из яда змеи рода Bungarus). По агонистам различают три типа рецепторов глутамата: квисквалатные(AMPA-тип), каинатные и NMDA (N-метил-D-аспартат) -типа. Некоторые рецепторы медиаторов (в частности, адренорецепторы и рецепторы многих нейропептидов) связаны не с ионными каналами (ионотропные рецепторы) , а с мембранным ферментом (метаботропные рецепторы) , например, аденилатциклазой . Последняя, однократно активируемая медиатором, катализирует превращение множества молекул аденозинтрифосфорной кислоты (АТФ) циклический аденозинмонофосфат (цАМФ) - усилительный механизм. ЦАМФ, являясь вторичным посредником, активирует в клетке многие ферменты, в частности протеинкиназы и таким образом стимулирует клеточный метаболизм. Разрушается цАМФ фосфодиэстеразой. Аденилатциклазной системе аналогичны гуанилатциклазная система, система фосфолипазы С и т.д. (рис.14). Образование цГМФ из нециклической формы катализируется гуанилатциклазой , активность которой стимулирует окись азота. Молекулы последней образуются при дезаминировании аргинина и образовании цитруллина под действием синтазы окиси азота. Активность этого фермента в свою очередь регулируется комплексом Са 2+ -кальмодулин. Таким образом, например, глутаматные рецепторы, запускающие входящий ток ионов Са 2+ , управляет колебаниями концентрации цГМФ в цитоплазме нейронов. В головном мозге позвоночных активность синтазы окиси азота выявляется в мозжечке, четверохолмии, полосатом теле и обонятельной луковице.
Механизм работы химического синапса: при деполяризации пресинаптической терминали (вызванной ПД или искусственно) в нее из среды входят ионы Ca 2+ , которые стимулируют процесс экзоцитоза - опорожнениявезикул в синаптическую щель.
Представлены характерные синаптические и везикулярные белки, а также их предполагаемые рецепторы и функции. Постулируются раздельные участки везикулярной мембраны для заякоривания пузырьков на цитоскелете), прикрепления везикулярной мембраны к пресинаптической и высвобождения медиатора через образовавшуюся пору. Молекулярные механизмы прикрепления везикул к пресинаптической мембране и образования поры предположительно различны. Некоторые из белков являются мишенями нейротоксинов, изменяющих выброс медиатора. Например, структура везикулярных белков синаптобревинов (VAMPs) нарушается под действием столбнячного и ботулинического токсинов; яд паука латротоксин связывается с пресинаптическими мембранными белками нейрексинами и усиливает опорожнение везикул.
1. Синапсины - белки, ассоциированные с везикулами, которые предположительно связывают синаптические пузырьки с цитоскелетом нервного окончания. 2. Прикрепление, образование поры и опорожнение везикул осуществляются взаимодействиями (указаны стрелками) различных везикулярных и мембранных белков. Например, в образовании прикрепительного комплекса участвуют везикулярные белки (синаптотагмин и синаптобревины) и белки плазматической мембраны нервного окончания (синтаксины и нейрексины). 3. Какие белки - плазматические или везикулярные образуют пору слияния до сих пор не ясно. Предположительно, это - синаптофизин (имеющий и другие функции) и белок плазматической мембраны физофилин. 4. Rab-белки могут участвовать в транспорте везикул в клетке и в прикреплении их к мембране нервной клетки. Везикулярные транспортеры играют роль в аккумуляции нейромедиатора в синаптических пузырьках.
Одновременно Ca 2+ начинает удаляться из цитоплазмы несколькими путями: связывание с белками, захват митохондриями и работа активного транспорта. Выход медиатора зависит от деполяризации терминали и составляет около 100 - 200 везикул, каждая из которых содержит одну порцию (квант), соответствующую приблизительно 10 4 молекул. Молекулы медиатора диффундируют к постсинаптической мембране, где взаимодействует с рецепторами постсинаптической мембраны, регулирующими состояние ионных каналов. Эта регуляция может быть прямой (как, например, в нервно-мышечном соединении скелетных мышц позвоночных), так и включать активацию систем вторичных внутриклеточных медиаторов (G-белки, цАМФ).
В первом случае осуществляется передача быстрых пусковых сигналов, во втором - осуществляются более медленные длительные воздействия. Направление изменения потенциала постсинаптической мембраны (деполяризация или гиперполяризация) зависит главным образом от того, открытием каких каналов управляют постсинаптические рецепторы. Часть молекул медиатора может взаимодействовать с пресинаптическими рецепторами, что приводит к изменению МП нервной терминали и, соответственно, количества выделяемого медиатора (обратная связь). Синаптическая щель очищается от медиатора различными путями: дезактивация, гидролиз, обратный захват в пресинаптическое окончание, диффузия, захват глиальными клетками. Основная часть синаптической задержки - времени от прихода нервного импульса до развития постсинаптического ответа (0,2-0,5 мс) приходится на процесс секреции медиатора. Химический синапс обеспечивает передачу сигнала только от пресинаптического нейрона к постсинаптическому.
При частой ритмической стимуляции в химических синапсах наблюдается сначала усиление (облегчение) , а затем ослабление (депрессия) передачи, т.е. рост, а затем падение амплитуды постсинаптических потенциалов. Эти явления в основном определяются изменениями в пресинаптическом звене. Они имеют особое развитие в некоторых синапсах ЦНС, где выступают как факторы синаптической пластичности. Наличие различных типов рецепторов в постсинаптической мембране может обусловливать развитие этих явлений по отдельности, как, например, длительную потенциацию и длительную депрессию.
Химические тормозные синапсы . Эти синапсы по механизму передачи возбуждения сходны с синапсами возбуждающего действия. тормозных синапсах медиатор (например, глицин) взаимодействует с рецепторами субсинаптической мембраны и открывает в ней хлорные каналы, это приводит к движению ионов хлора по концентрационному градиенту внутрь клетки и развитию гиперполяризации на субсинаптической: мембране. Возникает так называемый тормозной постсинаптический потенциал (ТПСП).
Ранее полагали, что каждому медиатору соответствует специфическая реакция постсинаптической клетки - возбуждение или торможение в той или иной форме. В настоящее время установлено, что одному медиатору чаще всего соответствует не один, а несколько различных рецепторов. Например, ацетилхолин в нервно-мышечных синапсах скелетных мышц действует на Н-холинорецепторы (чувствительные к никотину), которые открывают широкие каналы для натрия (и калия), что порождает ВПСП (ПКП) В ваго-сердечных синапсах тот же ацетилхолин действует на М-холинорецепторы (чувствительные к мускарину), открывающие селективные каналы для ионов калия, поэтому здесь генерируется тормозной постсинаптический потенциал (ТПСП). Следовательно, возбуждающий или тормозной характер действия медиатора определяется свойствами субсинаптической мембраны (точнее, видом рецептора), а не самого медиатора.
Физиологические свойства химических синапсов . Синапсы с химической передачей возбуждения обладают рядом общих свойств:
Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами субсинаптической мембраны;
Передача возбуждения через синапсы осуществляется медленнее, чем по нервному волокну - синаптическая задержка;
Передача возбуждения осуществляется с помощью специальных химических посредников - медиаторов;
В синапсах происходит трансформация ритма возбуждения;
Синапсы обладают низкой лабильностью;
Синапсы обладают высокой утомляемостью;
Синапсы обладают высокой чувствительностью к химическим (в том числе и к фармакологическим) веществам.
Электрические синапсы возбуждающего действия . Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока.
Схема передачи возбуждения в электрическом синапсе: ток, вызванный пресинаптическим потенциалом действия, раздражает постсинаптическую мембрану, где возникает ВПСП и потенциал действия. Поперечные каналы объединяют клетки не только электрически, но и химически, так как они проходимы для многих низкомолекулярных соединений. Поэтому возбуждающие электрические синапсы с поперечными каналами формируются, как правило, между клетками одного типа (например, между клетками сердечной мышцы).
Общими свойствами возбуждающих электрических синапсов являются:
Быстродействие (значительно превосходит таковое в химических синапсах);
Слабость следовых эффектов при передаче возбуждения (в результате этого в них практически невозможна суммация последовательных сигналов);
Высокая надежность передачи возбуждения.
Возбуждающие электрические синапсы могут возникать при благоприятных условиях и исчезать при неблагоприятных. Например, при повреждении одной из контактирующих клеток ее электрические синапсы с другими клетками ликвидируются. Это свойство называется пластичностью. Электрические синапсы могут быть с односторонней и двусторонней передачей возбуждения.
Электрический тормозный синапс. Наряду с электрическими синапсами возбуждающего действия могут встречаться электрические тормозные синапсы. Тормозящее влияние возникает за счет действия тока, вызванного потенциалом действия пресинаптической мембраны. Пресинаптический потенциал вызывает значительную гиперполяризацию сегмента и гиперполяризующий ток мгновенно тормозит генерацию потенциала действия в начальном сегменте аксона.
В смешанных синапсах пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Трофические влияния, передаваемые через синапсы. Помимо передачи возбуждающих и тормозных сигналов, которые имеют функциональное значение, синапсы обеспечивают трофические (т.е. затрагивающие рост и дифференцировку) взаимодействия контактирующих клеток, реализуемые с помощью трофических факторов белковой природы, вероятно, также аккумулируемых в везикулах. Эти факторы обеспечивают метаболическое поддержание необходимой структуры и свойств этих клеток. Двусторонние трофические взаимодействия предполагают во всех синапсах, но изучены они главным образом в скелетных нервно-мышечных синапсах позвоночных. Денервация мышцы приводит к потере мышечными волокнами дифференцировки, достигнутой в онтогенезе.
Постсинаптические процессы. Изменения потенциала постсинаптической мембраны в результате активации синапса называют синаптическим потенциалом. Возбуждающий постсинптический потенциал (ВПСП) возникает в деполяризующих синапсах и обусловлен одновременным повышением проницаемости мембраны для ионов Na + и K + . Возникающие при этом токи противоположно направлены (натриевый - внутрь клетки, калиевый - наружу). МП смещается в сторону деполяризации до значения равного полусумме равновесных потенциалов E Na и E K . Тормозные постсинаптические потенциалы(ТПСП) представляют собой гиперполяризационные изменения МП (до-80-90 мВ) и обусловлены открытием каналов для ионов K + (которые покидают клетку), либо ионов Cl - (входящих в клетку), либо для тех и других ионов одновременно. Амплитуда синаптических потенциалов зависит от количества выделяющегося медиатора (числа квантов) и, таким образом, эти реакции являются градуальными в отличие от ПД. Это амплитудное кодирование частотного сигнала осуществляется в постсинаптическом нейроне за исключением его аксонной области, в которой происходит возврат к частотному кодированию, благодаря распространяющимся по аксонному волокну ПД.
Создаваемая ВПСП деполяризация может значительно превышать, достигать или оставаться ниже порога возбуждения (отмечен точками). Амплитуда гиперполяризации вследствие развития ТПСП зависит от частоты пресинаптических ПД и влияет на частоту фоновой активности постсинаптической клетки.
Т.к. направление ионных токов зависит от градиента электрохимического потенциала данного иона, то амплитуда и полярность синаптического сигнала изменяются с изменением МП. Его значение, при котором происходит изменение знака синаптического действия, называется потенциалом реверсии. Распространение синаптических потенциалов обусловлено только физическими свойствами мембраны клетки и поэтому происходит с затуханием (уменьшением амплитуды). Синаптические потенциалы, возникающие в разных синапсах, могут взаимодействовать между собой, подчиняясь правилам алгебраического суммирования. Деполяризация, вызванная суммацией ВПСП, приближает МП к пороговой для возбуждения величине. Напротив, наложение ТПСП отдаляет его. Степень деполяризации или гиперполяризации мембраны зависит от «противоборства» между ионными проводимостями и токами, активируемыми при ВПСП и ТПСП. Главную роль в интеграции приходящих в клетку сигналов выполняют особые участки нейрональной мембраны: узлы ветвления дендритного дерева, соматическая мембрана, аксонный холмик. Чаще всего на дендритной мембране локализуются возбуждающие синапсы, в то время как тормозные располагаются, как правило, на соматической мембране. Окончательная интеграция всех входов происходит на аксонном холмике. Поскольку постсинаптические процессы возникают на различном удалении от триггерной зоны и распространяются пассивно, то их вклад в интегративный выход будет зависеть от локализации синаптических контактов.
На это взаимодействие влияют геометрические взаимоотношения между возбуждающими и тормозными синапсами, расположенными в разных участках дендритов, а также особенности электротонического распространения тока по этим дендритам. При этом в постсинаптическом нейроне при генерации ПД могут возникать эффекты сложения и вычитания входных сигналов, имеющих одинаковые или различные (возбуждающую и тормозную) модальности.
Передача сигнала неимпульсирующими нейронами. Некоторые нервные клетки не генерируют в физиологических условиях ПД (эта типичная для нейронов способность проявляется после некоторых видов химического воздействия). Примером таких клеток у позвоночных могут служить фоторецепторы и клетки-зерна обонятельных луковиц. Однако у этих нейронов сохраняется закономерность: медиатор выделяется только при деполяризации пресинаптической клетки.
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Законы проведения возбуждения по нервным волокнам
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
Закономерности проведения местного и распространяющегося возбуждения
Электротонический потенциал (местное возбуждение)
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
Потенциал действия (распространяющееся возбуждение)
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые и безмиелиновые нервные волокна
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты (шванновские клетки) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
Механизм проведения возбуждения по безмиелиновым нервным волокнам
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Механизм проведения возбуждения по миелиновым нервным волокнам
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье.При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Классификация нервных волокон
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Диаметр (мкм) |
Миелинизация |
Скорость про-ведения (м/с) |
Функциональное назначение |
|
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов |
||||
Чувствительные волокна кожных рецепторов |
||||
Чувствительные волокна проприорецепторов |
||||
Чувствительные волокна терморецепторов, ноцицепторов |
||||
Преганглионарные волокна симпатической НС |
||||
отсутствует |
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов |
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.

