20.01.2022
Miyelinsiz ve miyelinli sinir lifleri. "Sinir lifi" kavramı
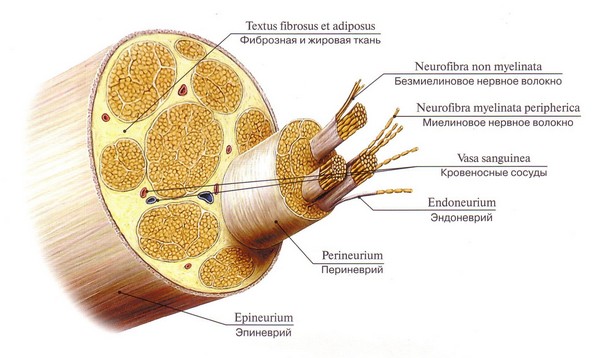
Sinir lifi, lemositlerle ve miyelinli veya miyelinsiz bir kılıfla kaplı uzun bir nöron sürecidir. Ana işlevi sinir uyarılarının iletilmesidir. Periferik ve merkezi sinir sisteminde, iskelet kaslarını innerve eden pulpa (miyelin) sinir lifleri baskındır; pulpasız olanlar ise sempatik departman otonom sistem ve iç organlara yayılır. Kılıfı olmayan liflere çıplak eksenel silindirler denir.
Sinir lifi, bir çeşit eksen oluşturan nöron sürecine dayanmaktadır. Dışarıda, nöron eksenine doğru spiral çizen çok sayıda mesakson dönüşünden oluşan biyomoleküler bir lipid bazlı miyelin kılıfı ile çevrilidir. Böylece sinir liflerinin miyelinasyonu meydana gelir.
Periferik sistemin miyelinli sinir lifleri ayrıca aksonu destekleyen ve nöron gövdesini besleyen yardımcı Schwann hücreleriyle kaplanır. Pulpa zarının yüzeyinde aralıklar vardır - Ranvier düğümleri, bu yerlerde eksenel silindir dış Schwann zarına bağlanır.
Miyelin tabakası elektriksel olarak iletken özelliklere sahip değildir; kesişmelerde bu özellikler vardır. Uyarma, dış uyaranın etki alanına en yakın Ranvier aralığında meydana gelir. Dürtü, bir müdahaleden diğerine spazmodik olarak iletilir, bu, yüksek bir dürtü yayılım hızı sağlar.
Miyelinli sinir lifleri kas dokusundaki metabolizmayı düzenler ve biyoelektrik akıma karşı yüksek dirence sahiptir.
Ranvier'in boşlukları dürtüleri üretir ve güçlendirir. Merkezi sinir sisteminin liflerinde Schwann zarı yoktur, bu fonksiyon oligodendroglia tarafından gerçekleştirilir.
Pulpasız dokular çok sayıda eksenel silindire sahiptir, miyelin tabakası ve kesişmeleri yoktur, üstleri Schwann hücreleriyle kaplıdır ve silindirlerle aralarında yarık benzeri boşluklar oluşur. Liflerin yalıtımı zayıftır, bir nöronun bir sürecinden diğerine bir impulsun yayılmasına izin verir ve bir nöronla temas halindedir. çevre Vücut daha fazla enerjiye ihtiyaç duyarken, dürtü iletim hızı kağıt hamuru liflerinden çok daha düşüktür.
Büyük sinir gövdeleri, nöronların pulpal ve pulpa dışı süreçlerinden oluşur; bunlar daha küçük demetlere ayrılır ve sinir uçlarıyla (reseptör, motor, sinapslar) biter.
Sinir uçları, miyelinli ve miyelinsiz sinir liflerinin internöron temaslarını, reseptör ve motor uçlarını oluşturan uçlarıdır.
Sınıflandırma ilkeleri

Farklı tipte sinir lifleri, uyarma impulslarının farklı iletim hızlarına sahiptir; bu, bunların çapına, aksiyon potansiyelinin süresine ve miyelinasyon derecesine bağlıdır. Hız ile lif çapı arasında doğru orantılı bir ilişki vardır.
Sinir liflerini sinir uyarılarının hızına göre sınıflandırmak için Erlanger-Gasser yapısal-fonksiyonel yöntemi:
- Miyelinli sinir lifi grubu A: α, β, Υ ve δ. En büyük çaplı ve en kalın kabuğun dokuları α - 20 μm'dir; iyi bir darbe iletim hızına sahiptirler - 120 m/sn. Bu dokular kolondaki uyarı kaynağını innerve eder. omurilik iskelet kası reseptörlerine, tendonlara ve dokunma duyularından sorumludur.
Diğer fiber türleri daha küçük bir çapa (12 μm) ve darbe iletim hızına sahiptir. Bu dokular ağrının kaynağı olan iç organlardan gelen sinyalleri merkezi sinir sistemine iletir.
- B grubu miyelin lifleri aittir. Toplam dürtü iletim hızı 14 m/sn'dir, aksiyon potansiyeli A grubu liflerinkinden 2 kat daha fazladır.Miyelin kılıfı iyi tanımlanmamıştır.
- C grubunun miyelinsiz lifleri çok küçük bir çapa (0,5 μ) ve uyarılma hızına (6 m/sn) sahiptir. Bu dokular sempatik sinir sistemini innerve eder. Bu grup aynı zamanda ağrı, soğuk, ısı ve basınç merkezlerinden gelen uyarıları ileten lifleri de içerir.
Nöronal süreçler afferent ve efferent olarak ikiye ayrılır. Birinci tip, impulsların doku reseptörlerinden merkezi sinir sistemine iletilmesini sağlar. İkinci tip uyarımı merkezi sinir sisteminden doku reseptörlerine iletir.
Afferent tip sinir liflerinin Lloyd-Hunt'a göre fonksiyonel sınıflandırması:
Demyenilizasyon
Sinir liflerinin demiyelinizasyon süreci, miyelin kılıfında doku fonksiyon bozukluğuna neden olan patolojik bir hasardır. Patolojiye inflamatuar süreçler, metabolik bozukluklar, nöroenfeksiyon, zehirlenme veya doku iskemisi neden olur. Miyelinin yerini fibröz plaklar alır, bunun sonucunda impulsların iletimi bozulur.
İlk demiyelinizasyon türü vücudun otoimmün reaksiyonlarının neden olduğu miyelinopati, Canavan hastalığı, Guillain-Barré sendromu, Charcot-Marie-Tooth amyotrofisidir.
İkinci tip miyelinoklastidir. Patoloji, miyelin kılıfının (Binswanger hastalığı) tahribatına kalıtsal bir yatkınlık ile karakterize edilir.
Demiyelinizan hastalıklar

Miyelin kılıfının tahrip olmasına yol açan hastalıklar çoğunlukla otoimmün niteliktedir; başka bir neden antipsikotiklerle tedavi veya kalıtsal yatkınlık olabilir. Lipid tabakasının tahribatı, tahriş dürtülerinin hızında bir azalmaya neden olur.
Hastalıklar merkezi sinir sistemini etkileyenlere ve periferik ağa zarar veren patolojilere ayrılır. Merkezi sinir sisteminin işleyişini etkileyen hastalıklar:
- Omuriliğin miyelopatisi, miyelin liflerinin sıkışması sonucu oluşur intervertebral fıtıklar, tümörler, kemik parçaları, sonra. Hastalarda etkilenen bölgedeki hassasiyet ve kas kuvveti azalır, kol veya bacaklarda felç oluşur, bağırsakların ve idrar sisteminin işleyişi bozulur, alt ekstremite kas atrofisi gelişir.
- Beynin lökodistrofisi hasara neden olur Beyaz madde. Hastaların hareket koordinasyonu zayıftır ve dengeyi sağlayamazlar. gelişen Kas Güçsüzlüğü istemsiz kasılmalar ve sinir tikleri ortaya çıkar. Bellek, entelektüel yetenekler, görme ve işitme giderek kötüleşir. Açık geç aşamalar körlük, sağırlık, tam felç ve yiyecekleri yutma güçlüğü ortaya çıkar.
- Beynin küçük fokal lökoensefalopatisi çoğunlukla 60 yaş üstü erkekleri etkiler. Başlıca nedenler: arteriyel hipertansiyon ve kalıtsal yatkınlık. Hastalarda hafıza ve dikkatte bozulma, uyuşukluk ve konuşma güçlüğü görülür. Yürüyüş yavaşlar, hareketlerin koordinasyonu bozulur, idrar kaçırma ortaya çıkar ve hasta yiyecekleri yutmakta zorluk çeker.
- Ozmotik demiyelinizasyon sendromu, beyin dokusundaki miyelin kılıflarının parçalanmasıyla karakterizedir. Hastalarda konuşma aparatında bir bozukluk, sürekli bir uyuşukluk hissi, depresyon veya artan uyarılabilirlik, mutizm, tüm ekstremitelerde parezi vardır. Açık erken aşamalar hastalıkta demiyelinizasyon süreci geri dönüşümlüdür.
- Multipl skleroz, bir veya iki uzuvda kısmi veya kısmi uyuşukluk ile kendini gösterir. toplam kayıp görme, gözleri hareket ettirirken ağrı, baş dönmesi, çabuk yorulma, uzuvlarda titreme, hareketlerin koordinasyon kaybı, vücudun çeşitli yerlerinde karıncalanma.
- Devic hastalığı, optik siniri ve omuriliği etkileyen inflamatuar bir otoimmün hastalıktır. Semptomlar körlük, paraparezi, tetraparezi ve pelvik organların işlev bozukluğunu içeren değişen derecelerde görme bozukluğunu içerir.
Hastalığın belirtileri miyelin liflerinin hasar alanına bağlıdır. Demiyelinizasyon süreci kullanılarak tespit edilebilir bilgisayarlı tomografi, manyetik rezonans terapisi. Elektromiyografide periferik sinir sisteminde hasar belirtileri tespit edilir.
Afferent görsel yolların miyelinasyonu Gebeliğin beşinci ayında lateral genikülat cisimde başlar ve doğumda kribriform plakta biter. CNS liflerinin miyelinlenmesinden sorumlu olan oligodendrositler normalde retinada yoktur. Histolojik çalışmalar, miyelinli sinir lifi bölgelerinde varsayılan oligodendrositlerin ve miyelinin varlığını ve bu bölgelerin dışında bulunmadığını doğruladı.
Şu tarihte: miyelinli liflerin diseksiyonu Rutin muayene sırasında gözlerin yaklaşık %1'inde, göz hastalarının ise %0,3-0,6'sında retinit saptanır.
Miyelinli sinir lifleri Genellikle diskin üst ve alt kutuplarında uzunlamasına çizgili demetler gibi görünürler. Ayrıca diskin değişen kısımlarının belirginleşmesi ve diskin kenarları ile alttaki liflerin üst üste binmesi nedeniyle kan damarları anormallik optik diskin şişmesini simüle edebilir.
Distalde lifler oluşur düzensiz yelpaze şeklindeki şekil. Miyelinasyon bölgesi içerisinde bazen küçük çatlaklar veya normal fundus alanları görülebilir. Vakaların %17-20'sinde iki taraflı miyelinli sinir lifleri tespit edilir. Vakaların %19'unda optik diske temas etmezler. Bazen, optik diskin nazal tarafındaki retinanın çevresinde izole edilmiş miyelinli sinir lifi demetleri tespit edilir.
Miyelin liflerinin patogenezi belirsizliğini koruyor, ancak lamina kribrozası zayıf olan veya bulunmayan hayvanlarda genellikle derin fizyolojik kazı ve retina sinir liflerinde yaygın miyelinasyon bulunurken, iyi gelişmiş lamina kribrosaya sahip hayvanlarda tamamen düz bir optik disk bulunur ve retinal miyelinasyon yoktur; bu da birkaç olası durumu akla getirir Patogenez mekanizmaları:
1. Lamina cribrosa'da, oligodendrositlerin retinaya nüfuz etmesine ve miyelin üretmesine izin veren bir kusurun varlığı.
2. Belki de akson sayısı skleral kanalın boyutuna göre küçüktür ve oküler kavitede miyelinasyonun devam etmesi için yeterli alan vardır. Periferik olarak izole edilmiş miyelinli sinir lifi demetlerinin bulunduğu gözlerde, lamina cribrosa oluşumunun zamanlamasının bozulması, oligodendrositlerin retinaya girmesine ve miyelinasyonun devam ettiği nispeten düşük sinir lifi yoğunluğuna sahip bir alana ulaşana kadar sinir lifi tabakası boyunca göç etmesine izin verir.
3. Daha sonra gelişme Kribriform plaka, oligodendrositlerin göz dokusuna göçü için koşullar yaratabilir.
Sinir liflerinin yaygın tek taraflı (veya nadiren iki taraflı) miyelinasyonuna miyopi eşlik edebilir yüksek derece ve kırılma ambliyopisi. Bu tür hastalarda miyelin örtüşür en tamamen olmasa da diskin çevresi. Maküler bölge (miyelin içermemesine rağmen) genellikle zayıflamış refleksler veya pigment dispersiyonu gibi anormallikler de gösterir. Makula bölgesinin durumu muhtemelen oklüzyon tedavisinin etkinliğinin en doğru göstergesidir.
Miyelinli sinir lifleri Gorlin sendromu (multipl bazal hücreli nevüs) ve otozomal dominant vitreoretinopatinin yanı sıra konjenital görme bozukluğu, retina sinir lifi tabakasında iki taraflı yaygın miyelinasyon, ciddi vitreus dejenerasyonu, yüksek miyopi, retina distrofisi, gece körlüğü ve uzuv deformiteleri eşlik edebilir.
Miyelinli sinir lifleri otozomal dominant bir şekilde kalıtsal olabilir. Anormal optik sinir uzunluğu (oksisefali), lamina kribrosa defektleri (eğik optik disk), ön segment disgenezisi ve nörofibromatozis tip 2 ile birlikte miyelinli sinir liflerinin izole vakaları tanımlanmıştır. Her ne kadar miyelinli sinir lifleri nörofibromatozise eşlik edebilse de, birçok yazar bu kombinasyonun şüpheli olduğunu düşünmektedir.
Bazen bölgeler sinir liflerinin miyelinasyonu Bebeklik döneminden sonra ve hatta yetişkinlikte bile ortaya çıkabilir. Bu gibi durumlarda bu anomalinin olağan nedeninin göz travması olması muhtemeldir ( künt travma bir vakada gözler ve diğerinde optik sinir kılıfının fenestrasyonu).
Belki böyle bir hasarla cribriform plakada bir kusur meydana gelir Oligodendrositlerin retinaya nüfuz etmesi için yeterlidir. Aksonlar hasar gördüğünde miyelinli sinir lifleri kaybolabilir.
Miyelinli sinir lifleri ile tek taraflı yüksek miyopi. Miyelinli sinir lifleri.
Miyelinli sinir lifleri. A. Hafif derece. B. Şiddetli derece.
Nöronların süreçleri neredeyse her zaman bir kılıfla (miyelin) kaplıdır. Bunun istisnası, bazı süreçlerin serbest sonlarıdır. Kılıfla birlikte olan bu işleme “sinir lifi” adı verilir.
Sinir lifi aşağıdakilerden oluşur: Eksenel silindir– bir sinir hücresinin süreci: akson veya dendrit
Glial kılıf eksenel silindiri bir kaplin şeklinde çevreler. CNS'de oligodendroglia tarafından ve PNS'de Schwann hücreleri tarafından oluşturulur (nörolemmositler bir tür oligodendrogliadır).
Sinir lifleri miyelinsiz ve miyelinli (miyelin kılıfına sahip) olarak sınıflandırılır.
Miyelinsiz sinir lifleri otonom sinir sisteminin bir parçasıdır ve efektör nöronların aksonları tarafından temsil edilir. Merkezi sinir sisteminde de bulunurlar ancak daha küçük miktarlardadırlar.
Yapı: Merkezde oligodendrositin (lemosit) çekirdeği bulunur ve çevre boyunca 10-20 eksenel silindir sitoplazmasına nüfuz eder. Bu tür sinir liflerine “kablo tipi lifler” de denir. Eksenel silindir oligodendrositin sitoplazmasına daldırıldığında, ikincisinin plazmalemmasının bölümleri birbirine yaklaşır ve bir mezenter oluşur - bir "mesakson" veya bir çift membran. Yüzeyde sinir lifi bir bazal membranla kaplıdır.
Miyelinli sinir lifleri merkezi sinir sisteminin, PNS'nin somatik kısımlarının ve otonom sinir sisteminin preganglionik kısımlarının bir parçasıdır. Hem aksonları hem de nöronların dendritlerini içerebilirler.
Yapı: Eksenel silindir her zaman 1'dir ve merkezde bulunur. Membranın 2 katmanı vardır: Schwann hücresinin çekirdeği ve sitoplazması ile temsil edilen iç (miyelin) ve dış (nörolemma). Dış tarafta bir bodrum zarı vardır. Miyelin tabakası oligodendrosit (lemosit) zarının bir parçasıdır. Membran eksenel bir silindir etrafında eşmerkezli olarak bükülür. Aslında bu çok uzun bir mesaksondur. Mesaksonlar dil benzeri sitoplazmik süreçler oluşturur.
Miyelinizasyon süreci miyelin kılıfının oluşumudur. Embriyogenezin geç evrelerinde ve doğumdan sonraki ilk aylarda ortaya çıkar.
CNS'de miyelinasyonun özelliklerinin bulunduğunu belirtmekte fayda var: 1 oligodendrosit, birkaç eksenel silindirin etrafında (dönen birkaç işlem kullanarak) bir miyelin kılıfı oluşturur. Bodrum zarı yok.
Miyelin lifinin yapısı.
Miyelin, Ranvier düğümlerinde düzenli olarak kesintiye uğrar. Yakalamalar arasındaki mesafe 0,3 – 1,5 nm'dir. Kesişme alanında eksenel silindirin trofizmi meydana gelir. Miyelinin yüzeyinde çentikler vardır. Bu miyelin diseksiyon alanları sinir lifinin esnekliğini arttırır ve esneme için bir “yedek” sağlar. Merkezi sinir sisteminde çentik yoktur.
Miyelin, lipitler için boyalarla boyanır: Sudan, Osmik asit.
Miyelin fonksiyonları:
Sinir uyarılarının iletim hızını arttırmak. Miyelinsiz liflerin hızı 1-2 m/s, miyelinli liflerin hızı ise 5-120 m/s'dir.
Na kanalları, biyoelektrik akımların ortaya çıktığı kesişme alanında yoğunlaşmıştır. Bir müdahaleden diğerine atlıyorlar. Bu sıçramalı iletimdir, yani bir dürtünün sıçramalarda iletilmesidir.
Miyelin, etrafa yayılan akımların girişini sınırlayan bir yalıtkandır.
Miyelinli ve miyelinsiz liflerin yapısındaki farklılıklar.
Miyelinsiz lif Miyelinli lif
Çoklu aks silindirleri 1 aks silindiri
Eksenel silindirler - aksonlar Eksenel silindirler bunlar ve diğerleri olabilir Eksenel silindirler miyelinsiz liflerden daha kalındır
Oligodendrosit çekirdeği merkezde, oligodendrosit çekirdeği ve sitoplazması lifin çevresinde bulunur.
Mezaxons kısadır Mezaxons eksenel silindir etrafında tekrar tekrar bükülür, bir miyelin kılıfı oluşur
Eksenel silindirin tüm uzunluğu boyunca Na kanalları Na kanalları yalnızca Ranvier düğümlerinde
Periferik sinirin yapısı.
Sinir, demetler halinde gruplandırılmış miyelinli ve miyelinsiz liflerden oluşur. Hem afferent hem de efferent lifleri içerir.
Sinir uyarılarının iletim mekanizmaları.
Sinapslar, bir sinyali bir hücreden diğerine aktarmak için kullanılan özel hücreler arası bağlantılardır.
Nöronların temas alanları birbirine çok yakındır. Ancak yine de çoğu zaman onları ayıran bir sinaptik yarık kalır. Sinaptik yarığın genişliği birkaç on nanometre mertebesindedir.
Nötronların başarılı bir şekilde çalışabilmesi için birbirlerinden izolasyonlarının sağlanması ve aralarındaki etkileşimin sinapslar tarafından sağlanması gerekir.
Sinapslar, yolları boyunca sinir sinyallerinin yükselticileri olarak görev yapar. Bu etki, nispeten düşük güçlü bir elektrik darbesinin daha önce birçok sinaptik kesecikte bulunan yüzbinlerce verici molekülü serbest bırakmasıyla elde edilir. Verici moleküllerden oluşan bir yaylım ateşi, kontrollü nöronun, postsinaptik reseptörlerin yoğunlaştığı küçük bir alanı üzerinde eşzamanlı olarak etki eder - sinyali şimdi kimyasal bir formdan elektriksel bir forma dönüştüren özel proteinler.
Şu anda arabulucunun serbest bırakılması sürecinin ana aşamaları iyi bilinmektedir. Bir sinir impulsu, yani bir elektrik sinyali, bir nöronda ortaya çıkar, süreçleri boyunca yayılır ve sinir uçlarına ulaşır. Kimyasal forma dönüşümü, durumu zarın elektrik alanı tarafından kontrol edilen presinaptik zardaki kalsiyum iyon kanallarının açılmasıyla başlar. Artık kalsiyum iyonları sinyal taşıyıcılarının rolünü üstleniyor. Açılan kanallardan sinir ucuna girerler. Keskin bir şekilde arttı Kısa bir zaman kalsiyum iyonlarının zara yakın konsantrasyonu, verici salınımı için moleküler makineyi harekete geçirir: sinaptik veziküller, dış zar ile daha sonra füzyon yapacakları bölgelere yönlendirilir ve son olarak içeriklerini sinaptik yarık boşluğuna bırakır.
Sinaptik iletim, uzaysal olarak ayrılmış iki sürecin dizisiyle gerçekleştirilir: sinaptik yarığın bir tarafında presinaptik ve diğer tarafında postsinaptik (Şekil 3). Kontrol nöronunun işlemlerinin uçları, aldıkları elektrik sinyallerine uyarak, sinaptik yarık boşluğuna özel bir aracı madde (verici) salar. Verici moleküller sinaptik yarıktan yeterince hızlı bir şekilde yayılır ve kontrol edilen hücrede (başka bir nöron, kas lifi, iç organların bazı hücreleri) bir yanıt elektrik sinyalini uyarır. Yaklaşık bir düzine farklı düşük moleküllü madde aracı olarak görev yapar:
asetilkolin (amino alkol kolin ve asetik asitin esteri); glutamat (glutamik asit anyonu); GABA (gama-aminobutirik asit); serotonin (triptofan amino asidinin türevi); adenozin vb.
Bunlar, mevcut ve nispeten ucuz hammaddelerden presinaptik nöron tarafından önceden sentezlenir ve vericinin özdeş kısımlarının (bir kesecikte birkaç bin molekül) sanki kaplardaymış gibi bulunduğu sinaptik keseciklerde kullanılıncaya kadar depolanır.
Sinaps diyagramı
En üstte, içine presinaptik reseptörlerin gömülü olduğu presinaptik membranla sınırlanan sinir ucunun bir bölümü bulunur; sinir ucunun içindeki sinaptik kesecikler bir vericiyle doludur ve onun serbest bırakılması için değişen derecelerde hazırdır; vezikül membranları ve presinaptik membran, presinaptik proteinler içerir. Aşağıda, postsinaptik reseptörlerin oluşturulduğu postsinaptik membranda kontrollü bir hücrenin bir bölümü bulunmaktadır.
Sinapslar bilgi akışını düzenlemek için uygun bir nesnedir. Bir sinaps yoluyla iletildiğinde sinyal amplifikasyonunun seviyesi, salınan aracı miktarı değiştirilerek, bilgi aktarımının tamamen yasaklanmasına kadar kolayca artırılabilir veya azaltılabilir. Teorik olarak bu, arabulucu salınımının herhangi bir aşamasını hedefleyerek yapılabilir.
Miyelin lifleri. Bazı sinir lifleri embriyogenez sırasında miyelinasyona uğrar: lemositler (Schwann hücreleri) önce aksona dokunur ve sonra onu sarar. Lemosit zarı akson etrafına bir rulo gibi sarılarak çok katmanlı bir spiral (miyelin kılıfı) oluşturur. Miyelin kılıfı sürekli değildir - sinir lifinin tüm uzunluğu boyunca, birbirinden eşit mesafelerde küçük kırılmalar (Ranvier düğümleri) vardır. Engelleme alanında akson miyelin kılıfından yoksundur.
Miyelinsiz lifler. Diğer liflerin miyelinasyonu erken aşamalarda sona erer embriyonik gelişme. Bir veya daha fazla akson lemositin içine nüfuz eder; onları tamamen veya kısmen çevreler ancak çok katmanlı bir miyelin kılıfı oluşturmaz.
Miyelinsiz sinir lifleri boyunca uyarılma mekanizması
Dinlenme halinde, sinir lifi zarının tüm iç yüzeyi negatif yük taşır ve zarın dış tarafı pozitif yük taşır. Membranın iç ve dış kısımları arasında elektrik akımı akmaz çünkü Lipid zarı yüksek elektrik direncine sahiptir.Bir aksiyon potansiyelinin gelişimi sırasında, zarın uyarılmış kısmında yük tersinmesi meydana gelir. Uyarılmış ve uyarılmamış alanların sınırında bir elektrik akımı akmaya başlar. Elektrik akımı, zarın en yakın bölümünü tahriş eder ve onu uyarılma durumuna getirirken, önceden uyarılan alanlar dinlenme durumuna geri döner. Böylece uyarılma dalgası sinir lifi zarının tüm yeni alanlarını kapsar.
Miyelinli sinir lifleri boyunca uyarılma mekanizması
Miyelinli bir sinir lifinde, miyelin kılıfıyla kaplanan membran kısımları uyarılamaz; uyarma yalnızca Ranvier düğümleri bölgesinde bulunan membran alanlarında meydana gelebilir.Ranvier düğümlerinden birinde AP'nin gelişmesiyle birlikte, membran yükünün tersine çevrilmesi meydana gelir. Membranın elektronegatif ve elektropozitif alanları arasında, zarın komşu bölgelerini tahriş eden bir elektrik akımı ortaya çıkar. Bununla birlikte, zarın yalnızca bir sonraki Ranvier düğümü bölgesindeki kısmı uyarılma durumuna geçebilir. Böylece uyarılma, membran boyunca spazmodik (tuzlayıcı) bir şekilde bir Ranvier düğümünden diğerine yayılır.
NÖRON FONKSİYONLARI
Merkezi sinir sistemini ve iletkenlerini inşa etmek için kullanılan malzeme, sinir hücreleri (nöronlar) ve nöroglia olmak üzere iki bileşenden oluşan sinir dokusudur. Merkezi sinir sisteminin ana işlevsel unsurları nöronlardır: Hayvan vücudunda yaklaşık 50 milyar nöron vardır ve bunların yalnızca küçük bir kısmı vücudun çevresel kısımlarında bulunur.
Nöronlar toplam sayının %10 - 15'ini oluşturur hücresel elementler sinir sisteminde. Ana kısmı nöroglial hücreler tarafından işgal edilmiştir. Yüksek hayvanlarda farklılaşmış nöronlar doğum sonrası intogenez sırasında bölünmez. (daha önce düşünüldüğü gibi). Nöronlar şekil (piramidal, yuvarlak, yıldız şeklinde, oval), boyut (5 ila 150 µm arası) ve işlem sayısı bakımından önemli ölçüde farklılık gösterir, ancak aynı zamanda ortak özelliklere de sahiptirler.
Beynin bazı kısımlarında mukoprotein veya glikoprotein yapısında salgı granülleri üreten nöronlar vardır. Aynı zamanda sahipler fizyolojik belirtiler nöronlar ve glandüler hücreler. Bu hücrelere nörosekretuar hücreler denir. Nöronların işlevi, reseptörlerden veya diğer sinir hücrelerinden gelen sinyalleri algılamak, bilgiyi depolamak ve işlemek ve sinir uyarılarını diğer hücrelere (sinir, kas veya salgı) iletmektir. Buna göre nöronların uzmanlaşması gerçekleşir. Bunlar 3 gruba ayrılır: hassas (duyusal, afferent) nöronlar, asosiyatif (ara, interkalar) nöronlar, motor (efektör) nöronlar.
Nöroglia. Toplu sinir dokusu yardımcı işlevleri yerine getiren ve nöronlar arasındaki boşluğun neredeyse tamamını dolduran glial elemanları oluşturur. Anatomik olarak beyindeki nöroglial hücreler (oligodendrositler ve astrositler) ile periferik sinir sistemindeki Schwann hücreleri arasında ayrım yapılır. Oligodendrositler ve Schwann hücreleri aksonların çevresinde miyelin kılıfları oluşturur. Glia hücreleri ve nöronlar arasında 15-20 nm genişliğinde, birbirleriyle iletişim kurarak sıvıyla dolu bir ara boşluk oluşturan boşluklar vardır. Bu boşluk sayesinde, nöron ve glial hücreler arasındaki madde alışverişinin yanı sıra nöronlara oksijen ve oksijen sağlanması da meydana gelir. besinler difüzyon yoluyla. Glial hücreler görünüşe göre sadece destek görevi görüyor ve koruyucu işlevler merkezi sinir sisteminde bulunur ve beklendiği gibi bir beslenme veya bilgi depolama kaynağı değildir.
Membran özellikleri açısından glial hücreler nöronlardan farklıdır: elektrik akımına pasif olarak tepki verirler, zarları yayılan bir dürtü oluşturmaz. Nöroglial hücreler arasında doğrudan elektriksel iletişimi sağlayan sıkı bağlantılar (düşük dirençli alanlar) bulunur. Glial hücrelerin membran potansiyeli nöronlarınkinden daha yüksektir ve esas olarak ortamdaki K+ iyonlarının konsantrasyonuna bağlıdır.
Hücre dışı boşluktaki nöronların aktif aktivitesi sırasında K+ konsantrasyonu arttığında, bunun bir kısmı depolarize glial elementler tarafından emilir. Glia'nın bu tamponlama fonksiyonu nispeten sabit bir hücre dışı K+ konsantrasyonu sağlar. Glial hücreler - astrositler - nöronların gövdeleri ile kılcal damarların duvarı arasında bulunur, süreçleri ikincisinin duvarı ile temas halindedir. Bu perivasküler süreçler kan-beyin bariyerinin unsurlarıdır. Mikroglial hücreler fagositik bir işlev görür; beyin dokusu hasar gördüğünde sayıları keskin bir şekilde artar.
Sinapsların fizyolojisi.
Kimyasal sinapslar nispeten geniş bir sinaptik yarığa (20 - 50 nm) ve sinaptik membranların yüksek direncine sahiptir. Presinaptik sinir terminali çok sayıda vezikül içerir. Sinaptik veziküller - yaklaşık 50 nm çapında, bir aracı madde ile doldurulmuş.
Kimyasal uyarıcı sinapslarda uyarılma aktarım mekanizması. Kimyasal iletimli sinapslarda uyarım kullanılarak iletilir. arabulucular(aracılar). Mediat ory- Bu kimyasal maddeler Sinapslarda uyarımın iletilmesini sağlar. Niteliklerine bağlı olarak arabulucular birkaç gruba ayrılır:
monoaminler(asetilkolin, dopamin, norepinefrin, serotonin, vb.);
Amino asitler(gama-aminobutirik asit - GABA, glutamik asit, glisin, vb.);
nöropeptidler(P maddesi, endorfinler, nörotensin, ACTH, anjiyotensin, vazopressin, somatostatin, vb.). Moleküler formdaki verici, girdiği presinaptik kalınlaşmanın (sinaptik plak) keseciklerinde bulunur:
Hızlı aksonal taşıma (aksocurrent) kullanılarak nöronun perinükleer bölgesinden;
Sinaptik terminallerde bölünme ürünlerinden meydana gelen aracının sentezi nedeniyle;
Vericinin sinaptik yarıktan değişmeden geri alınması nedeniyle.
Çeşitli aracı sistemler. Dale'in ilkesi: bir nöron, kural olarak, tüm terminallerinde bir nörotransmitteri sentezler ve kullanır. Bir nöronun birden fazla verici kullanması mümkündür (komedyenler) , ama görünüşe göre aynı kombinasyonda (bu prensip şu anda tartışmalıdır, arabulucu sistemler hakkında aşağıya bakınız) . Dale ilkesinin bir sonucu olarak aşağıdaki hükümler dikkate alınabilir:
1) Sinaptik etkinin işareti verici tarafından değil, postsinaptik hücre üzerindeki reseptörlerin özellikleri tarafından belirlenir.
2) Aynı presinaptik nörona postsinaptik olan hücreler üzerindeki reseptörler, farmakolojik olarak farklı olabilir ve farklı iyon kanallarını kontrol edebilir.
3) Bir postsinaptik hücre, belirli bir verici için birden fazla tipte reseptöre sahip olabilir ve bu reseptörlerin her biri, farklı bir iyon iletimi mekanizmasını kontrol edebilir.
Bu üç özelliğin bir sonucu olarak, hücreler hem farklı postsinaptik hücreler hem de aynı hücre üzerinde zıt sinaptik eylemler uygulayabilir.
Bugüne kadar hayvanlarda ve insanlarda tanımlanan aracılar oldukça heterojen bir madde grubu oluşturur. Monoaminler: asetilkolin, dopamin, norepinefrin, serotonin (5-hidroksitriptamin, 5-HT), histamin. Amino asitler: gama-aminobütirik asit (GABA), glutamik asit, glisin, taurin vb. Nöropeptitler: metenkefalin, lehenkefalin, endorfin, oksitosin ,vazopressin ve benzeri . İlgili sinapslara örneğin şu ad verilir: kolinerjik, serotonerjik, noradrenerjik vesaire. Çok sayıda aracı, çok sayıda postsinaptik reseptöre karşılık gelir: kolinoreseptörler , adrenoreseptörler, GABA reseptörleri vesaire. Her reseptör grubunda çeşitli alt tipler bulunur; örneğin nikotinik ve muskarinik kolinerjik reseptörler (sırasıyla N-ChR ve M-ChR).
Reseptörlerin ayrılması farklılıklarına dayanmaktadır. farmakolojik özellikler: farklı agonistler (bir aracının etkisini taklit eden maddeler) ve düşmanlar (arabulucunun etkisine müdahale eden maddeler). Örneğin H-ChR için agonist nikotindir , antagonistleri - tubocurarine (kürar zehirinden izole edilmiştir), bungarotoksin (Bungarus cinsinin yılan zehirinden izole edilmiştir). Agonistlere göre üç tip glutamat reseptörü vardır: kisqualate (AMPA tipi), kainat Ve NMDA (N-metil-D-aspartat) -tip. Bazı nörotransmiter reseptörleri (özellikle adrenoreseptörler ve birçok nöropeptit reseptörü) iyon kanallarıyla ilişkili değildir. (iyonotropik reseptörler) ve bir membran enzimi ile (metabotropik reseptörler) , Örneğin, adenilat siklaz . İkincisi, bir aracı tarafından aktive edildiğinde birçok molekülün dönüşümünü katalize eder. adenosin trifosforik asit (ATP) döngüsel adenozin monofosfat (cAMP) - amplifikatör mekanizması. İkincil haberci olan CAMP, hücrede özellikle birçok enzimi aktive eder. protein kinazlar ve böylece hücresel metabolizmayı uyarır. cAMP yok edildi fosfodiesteraz. Adenilat siklaz sistemi guanilat siklaz sistemine, fosfolipaz C sistemine vb. benzer. (Şekil 14). Siklik olmayan formdan cGMP'nin oluşumu aşağıdakiler tarafından katalize edilir: guanilat siklaz aktivitesi nitrik oksit tarafından uyarılır. İkincisinin molekülleri, argininin deaminasyonu ve sitrülinin etkisi altında oluşumu sırasında oluşur. nitrik oksit sentezi. Bu enzimin aktivitesi de kompleks tarafından düzenlenir. Ca2+ -kalmodulin. Böylece, örneğin Ca2+ iyonlarının gelen akımını tetikleyen glutamat reseptörleri, nöronların sitoplazmasındaki cGMP konsantrasyonundaki dalgalanmaları kontrol eder. Omurgalı beyninde, beyincik, kuadrigeminal bölge, striatum ve koku alma soğanında nitrik oksit sentaz aktivitesi tespit edilir.
Kimyasal sinapsın mekanizması: presinaptik terminalin depolarizasyonu sırasında (AP'nin neden olduğu veya yapay olarak), Ca2+ iyonları çevreden girer ve bu da süreci uyarır ekzositoz - veziküllerin sinaptik yarığa boşaltılması.
Karakteristik sinaptik ve veziküler proteinler, varsayılan reseptörleri ve fonksiyonlarıyla birlikte sunulmaktadır. Veziküllerin hücre iskeletine sabitlenmesi, veziküler membranın presinaptik membrana bağlanması ve vericinin ortaya çıkan gözenek yoluyla serbest bırakılması için veziküler membranın ayrı bölümlerinin olduğu varsayılmaktadır. Moleküler mekanizmalar veziküllerin presinaptik membrana bağlanması ve bir gözenek oluşumu muhtemelen farklıdır. Proteinlerden bazıları, nörotransmitter salınımını değiştiren nörotoksinlerin hedefleridir. Örneğin veziküler sinaptobrevin proteinlerinin (VAMP'ler) yapısı tetanoz ve botulinum toksinleri tarafından bozulur; Örümcek zehiri latrotoksin, presinaptik membran proteinleri olan neurexinlere bağlanır ve veziküler boşalmayı arttırır.
1. Sinapsinler, muhtemelen sinaptik kesecikleri sinir terminalinin hücre iskeletine bağlayan keseciklerle ilişkili proteinlerdir. 2. Bağlanma, gözenek oluşumu ve veziküllerin boşaltılması, çeşitli veziküler ve membran proteinlerinin etkileşimleri (oklarla gösterilen) ile gerçekleştirilir. Örneğin, veziküler proteinler (sinaptotagmin ve sinaptobrevinler) ve sinir ucunun plazma zarının proteinleri (sözditaksinler ve neureksinler) bağlanma kompleksinin oluşumuna katılır. 3. Füzyon gözeneklerini hangi proteinlerin (plazmatik veya veziküler) oluşturduğu henüz belli değil. Muhtemelen bunlar sinaptofizin (başka işlevlere de sahiptir) ve plazma zarı proteini fizofilindir. 4. Rab proteinleri hücre içindeki keseciklerin taşınmasına ve bunların sinir hücresi zarına bağlanmasına katılabilir. Vesiküler taşıyıcılar, sinaptik veziküllerde nörotransmitter birikiminde rol oynar.
Aynı zamanda Ca2+ sitoplazmadan çeşitli yollarla uzaklaştırılmaya başlar: proteinlere bağlanma, mitokondri tarafından alım ve aktif taşıma. Aracının salınması, terminalin depolarizasyonuna bağlıdır ve yaklaşık 100 - 200 keseciktir; bunların her biri, yaklaşık 104 moleküle karşılık gelen bir kısım (kuantum) içerir. Aracı moleküller postsinaptik membrana yayılır ve burada iyon kanallarının durumunu düzenleyen postsinaptik membran reseptörleri ile etkileşime girerler. Bu düzenleme doğrudan olabilir (örneğin omurgalı iskelet kaslarının nöromüsküler kavşağında olduğu gibi) veya sistemlerin aktivasyonunu içerebilir. ikincil hücre içi aracılar (G proteinleri, cAMP).
İlk durumda hızlı tetikleme sinyalleri iletilir, ikincisinde ise daha yavaş, uzun vadeli etkiler gerçekleştirilir. Postsinaptik membran potansiyelindeki değişimin yönü (depolarizasyon veya hiperpolarizasyon), esas olarak hangi kanalların açılmasının postsinaptik reseptörler tarafından kontrol edildiğine bağlıdır. Bazı aracı moleküller etkileşime girebilir presinaptik reseptörler, bu, sinir terminalinin MP'sinde ve buna bağlı olarak salınan aracı madde miktarında (geribildirim) bir değişikliğe yol açar. Sinaptik yarık, vericiden çeşitli yollarla temizlenir: deaktivasyon, hidroliz, presinaptik terminale yeniden alım, difüzyon ve glial hücreler tarafından alım. Ana bölüm sinaptik gecikme - Sinir impulsunun gelmesinden postsinaptik yanıtın gelişmesine kadar geçen süre (0,2-0,5 ms), vericinin salgılanması süreciyle açıklanır. Kimyasal bir sinaps, yalnızca presinaptik nörondan postsinaptik nörona sinyal iletimini sağlar.
Kimyasal sinapslarda sık ritmik uyarımla birlikte artış olur. (rahatlama) ve ardından zayıflama (depresyon) iletim, yani postsinaptik potansiyellerin genliğinde bir artış ve ardından bir azalma. Bu fenomenler esas olarak presinaptik bağlantıdaki değişikliklerle belirlenir. Merkezi sinir sisteminin bazı sinapslarında özel bir gelişimleri vardır ve burada sinaptik plastisite faktörleri olarak hareket ederler. Postsinaptik membranda farklı tipteki reseptörlerin varlığı, uzun vadeli güçlenme ve uzun vadeli depresyon gibi bu fenomenlerin gelişimini ayrı ayrı belirleyebilir.
Kimyasal inhibitör sinapslar. Bu sinapslar, uyarıcı sinapslara uyarı aktarım mekanizmaları bakımından benzerdir. İnhibitör sinapslarda, bir aracı (örneğin, glisin) subsinaptik membranın reseptörleri ile etkileşime girer ve içindeki klorür kanallarını açar, bu, klor iyonlarının konsantrasyon gradyanı boyunca hücreye hareketine ve subsinaptikte hiperpolarizasyonun gelişmesine yol açar. zar. Sözde bir şey var inhibitör postsinaptik potansiyel(TPSP).
Daha önce, her aracının postsinaptik hücrenin belirli bir reaksiyonuna - şu veya bu şekilde uyarılma veya inhibisyon - karşılık geldiğine inanılıyordu. Artık bir vericinin çoğu zaman bir değil, birkaç farklı reseptöre karşılık geldiği tespit edilmiştir. Örneğin, iskelet kaslarının nöromüsküler sinapslarındaki asetilkolin, EPSP (EPSP) oluşturan sodyum (ve potasyum) için geniş kanallar açan H-kolinerjik reseptörler (nikotine duyarlı) üzerinde etki gösterir. Vago-kardiyak sinapslarda aynı asetilkolin etki eder. M-kolinerjik reseptörler (muskarine duyarlı) üzerinde potasyum iyonları için seçici kanallar açarak burada bir inhibitör postsinaptik potansiyel (IPSP) oluşturulur. Sonuç olarak, aracının etkisinin uyarıcı veya engelleyici doğası, aracının kendisi tarafından değil, subsinaptik zarın özellikleri (daha kesin olarak reseptör türü) tarafından belirlenir.
Fizyolojik özellikler kimyasal sinapslar. Uyarımın kimyasal iletimi ile sinapsların bir dizi ortak özelliği vardır:
Sinapslar aracılığıyla uyarılma yalnızca tek yönde (tek taraflı) gerçekleştirilir. Bunun nedeni sinapsın yapısından kaynaklanmaktadır: verici yalnızca presinaptik kalınlaşmadan salınır ve subsinaptik membranın reseptörleri ile etkileşime girer;
Uyarımın sinapslar yoluyla iletilmesi, sinir lifi - sinaptik gecikme yoluyla olduğundan daha yavaştır;
Uyarma aktarımı, özel kimyasal aracılar - aracılar yardımıyla gerçekleştirilir;
Sinapslarda uyarılma ritminde bir dönüşüm meydana gelir;
Sinapsların değişkenliği düşüktür;
Sinapslar oldukça yorulabilir;
Sinapslar kimyasal (farmakolojik dahil) maddelere karşı oldukça duyarlıdır.
Uyarıcı eylemin elektriksel sinapsları. Uyarma kimyasal iletimi olan sinapslara ek olarak, elektrik iletimi olan sinapslar esas olarak merkezi sinir sisteminde (CNS) bulunur. Uyarıcı elektriksel sinapslar, çok dar bir sinaptik yarık ve yerel elektrik akımlarının verimli geçişini sağlayan bitişik pre ve postsinaptik membranların çok düşük spesifik direnci ile karakterize edilir. Düşük direnç genellikle her iki membranı geçen, yani hücreden hücreye giden (boşluk kavşağı) enine kanalların varlığıyla ilişkilidir. Kanallar, temas eden zarların her birinin tamamlayıcı bir şekilde bağlanan protein molekülleri (yarı moleküller) tarafından oluşturulur. Bu yapı elektrik akımına kolaylıkla geçebilmektedir.
Elektriksel bir sinapsta uyarılma iletim şeması: Presinaptik aksiyon potansiyelinin neden olduğu akım, EPSP ve aksiyon potansiyelinin meydana geldiği postsinaptik membranı tahriş eder. Enine kanallar hücreleri yalnızca elektriksel olarak değil aynı zamanda kimyasal olarak da birbirine bağlar, çünkü bunlar birçok düşük moleküllü bileşik için kullanılabilirdir. Bu nedenle, kural olarak aynı tipteki hücreler arasında (örneğin kalp kası hücreleri arasında) enine kanallara sahip uyarıcı elektriksel sinapslar oluşur.
Uyarıcı elektriksel sinapsların genel özellikleri şunlardır:
Eylem hızı (kimyasal sinapslarınkini önemli ölçüde aşar);
Uyarma aktarımı sırasında iz etkilerinin zayıflığı (bunun sonucunda ardışık sinyallerin içlerinde toplanması neredeyse imkansızdır);
Uyarma iletiminin yüksek güvenilirliği.
Uyarıcı elektriksel sinapslar uygun koşullar altında ortaya çıkabilir ve olumsuz koşullar altında kaybolabilir. Örneğin temas halindeki hücrelerden biri hasar görürse diğer hücrelerle olan elektriksel sinapsları ortadan kalkar. Bu özelliğe plastisite denir. Elektriksel sinapslar tek yönlü veya iki yönlü uyarım iletimi ile olabilir.
Elektriksel inhibitör sinaps. Uyarıcı etkinin elektriksel sinapslarının yanı sıra, elektriksel inhibitör sinapslar da bulunabilir. İnhibitör etki, presinaptik membranın aksiyon potansiyelinin neden olduğu akımın etkisi nedeniyle ortaya çıkar. Presinaptik potansiyel, segmentin önemli ölçüde hiperpolarizasyonuna neden olur ve hiperpolarize edici akım, aksonun başlangıç segmentinde bir aksiyon potansiyelinin oluşmasını anında engeller.
İÇİNDE karışık sinapslar presinaptik aksiyon potansiyeli, pre- ve postsinaptik membranların birbirine sıkı bir şekilde bitişik olmadığı tipik bir kimyasal sinapsın postsinaptik membranını depolarize eden bir akım üretir. Dolayısıyla bu sinapslarda kimyasal iletim gerekli bir takviye mekanizması görevi görür.
Sinapslar yoluyla iletilen trofik etkiler. Sinapslar, işlevsel önemi olan uyarıcı ve engelleyici sinyallerin iletilmesinin yanı sıra, trofik (yani büyümeyi ve farklılaşmayı etkileyen) temas halindeki hücrelerin protein niteliğindeki trofik faktörlerin yardımıyla gerçekleştirilen etkileşimleri, muhtemelen keseciklerde de birikmiştir. Bu faktörler, bu hücrelerin gerekli yapı ve özelliklerinin metabolik olarak korunmasını sağlar. Tüm sinapslarda çift yönlü trofik etkileşimler öne sürülmüştür, ancak öncelikle omurgalı iskelet nöromüsküler sinapslarında incelenmiştir. Kas denervasyonu, intogenez sırasında elde edilen kas lifi farklılaşmasının kaybına yol açar.
Postsinaptik süreçler. Sinaps aktivasyonu sonucu postsinaptik membranın potansiyelindeki değişikliklere denir. sinaptik potansiyel. Uyarıcı postsinaptik potansiyel (EPSP) depolarize edici sinapslarda meydana gelir ve Na + ve K + iyonları için membran geçirgenliğinde eş zamanlı bir artıştan kaynaklanır. Bu durumda ortaya çıkan akımlar ters yöndedir (hücrenin içindeki sodyum, dışarıdaki potasyum). MF, E Na ve E K denge potansiyellerinin toplamının yarısına eşit bir değere depolarizasyona doğru kayar. İnhibitör postsinaptik potansiyeller (IPSP'ler) MF'deki (-80-90 mV'ye kadar) hiperpolarizasyon değişikliklerini temsil eder ve K + iyonları (hücreyi terk eden) veya Cl iyonları (hücreye giren) veya her iki iyon için kanalların açılmasından kaynaklanır. zaman. Sinaptik potansiyellerin genliği salınan aracının miktarına (kuantum sayısına) bağlıdır ve dolayısıyla bu reaksiyonlar kademeli PD'den farklı olarak. Frekans sinyalinin bu genlik kodlaması, akson lifi boyunca yayılan AP'ler nedeniyle frekans kodlamasına geri dönüşün meydana geldiği aksonal bölgesi haricinde postsinaptik nöronda meydana gelir.
EPSP tarafından oluşturulan depolarizasyon, uyarılma eşiğini (noktalarla gösterilir) önemli ölçüde aşabilir, ulaşabilir veya altında kalabilir. IPSP'nin gelişimine bağlı hiperpolarizasyonun genliği, presinaptik AP'lerin frekansına bağlıdır ve postsinaptik hücrenin arka plan aktivitesinin sıklığını etkiler.
Çünkü İyonik akımların yönü, belirli bir iyonun elektrokimyasal potansiyelinin gradyanına bağlıdır, daha sonra sinaptik sinyalin genliği ve polaritesi MF'deki değişikliklerle değişir. Sinaptik eylemin işaretinin değiştiği değere denir geri dönüş potansiyeli. Sinaptik potansiyellerin yayılması yalnızca belirlenir fiziki ozellikleri hücre zarı ve dolayısıyla zayıflama (genlikte azalma) ile ortaya çıkar. Farklı sinapslarda ortaya çıkan sinaptik potansiyeller, cebirsel toplama kurallarına uyarak birbirleriyle etkileşime girebilir. EPSP'lerin toplamının neden olduğu depolarizasyon, MP'yi uyarılma için eşik değerine yaklaştırır. Tam tersine, IPSP'nin dayatılması onu uzaklaştırıyor. Membranın depolarizasyon veya hiperpolarizasyon derecesi, iyonik iletkenlikler ile EPSP ve IPSP tarafından etkinleştirilen akımlar arasındaki "karşıtlığa" bağlıdır. Hücreye giren sinyallerin entegrasyonundaki ana rol, nöronal membranın özel alanları tarafından oynanır: dendritik ağacın dallanma düğümleri, somatik membran, akson tepeciği. Çoğu zaman, uyarıcı sinapslar dendritik membran üzerinde lokalize olurken, inhibitör sinapslar kural olarak somatik membran üzerinde bulunur. Tüm girdilerin son entegrasyonu akson tepeciğinde gerçekleşir. Postsinaptik süreçler tetikleme bölgesinden farklı uzaklıklarda ortaya çıktığı ve pasif olarak yayıldığından, bütünleştirici çıktıya katkıları sinaptik temasların lokalizasyonuna bağlı olacaktır.
Bu etkileşim, dendritlerin farklı kısımlarında bulunan uyarıcı ve engelleyici sinapslar arasındaki geometrik ilişkilerden ve ayrıca bu dendritler boyunca akımın elektrotonik yayılımının özelliklerinden etkilenir. Bu durumda postsinaptik nöronda AP'lerin üretimi sırasında aynı veya farklı (uyarıcı ve inhibitör) modalitelere sahip giriş sinyallerinin toplanması ve çıkarılmasının etkileri ortaya çıkabilir.
Darbeli olmayan nöronlar tarafından sinyal iletimi. Bazı sinir hücreleri, fizyolojik koşullar altında AP üretmez (nöronlara özgü olan bu yetenek, belirli kimyasal maruziyet türlerinden sonra ortaya çıkar). Omurgalılardaki bu tür hücrelerin örnekleri, fotoreseptörler ve koku soğanlarının granül hücreleridir. Bununla birlikte, bu nöronlar bir modeli korur: Verici yalnızca presinaptik hücre depolarize olduğunda serbest bırakılır.
Uyarılabilir bir hücrenin zarının bir kısmında ortaya çıkan uyarılma, yayılma yeteneğine sahiptir. Bir nöronun uzun uzantısı olan akson (sinir lifi), vücutta uzun mesafeler boyunca uyarılmanın iletilmesi gibi spesifik bir işlevi yerine getirir.
Uyarma yasalarısinir lifleri boyunca
Anatomik ve fizyolojik süreklilik yasası - uyarım, ancak morfolojik ve işlevsel olarak sağlamsa sinir lifi boyunca yayılabilir.
İki yönlü uyarım iletimi kanunu– Sinirin bir kısmında meydana gelen uyarılma, çıktığı yerden her iki yöne yayılır. Vücutta uyarılma her zaman hücre gövdesinden akson boyunca (ortodromik olarak) yayılır.
Yalıtılmış iletim kanunu– Sinirin bir parçası olan lif boyunca yayılan uyarım, komşu sinir liflerine iletilmez.
Yerel desenlerve heyecan yayılıyor
Elektrotonik potansiyel (yerel uyarım)
zayıflama ile sinir lifleri boyunca yayılır (ile azalma), yani. yerel tepkinin genliği, menşe yerinden uzaklaştıkça hızla azalır;
zayıflama nedeniyle yerel tepki kısa mesafelere yayılır (en fazla 2 cm);
yerel uyarılma, hücre enerjisini harcamadan pasif olarak yayılır;
yerel uyarımın yayılma mekanizması, iletkenlerdeki elektrik akımının yayılmasına benzer; uyarılmanın bu yayılma yöntemine denir elektrotonik.
Aksiyon potansiyeli (uyarılmanın yayılması)
sinir lifleri boyunca zayıflama olmadan yayılır, aksiyon potansiyelinin genliği, menşe yerinden herhangi bir mesafede aynıdır;
aksiyon potansiyelinin yayıldığı mesafe yalnızca sinir lifinin uzunluğuyla sınırlıdır;
aksiyon potansiyelinin yayılması, fiberin iyon kanallarının durumunun değiştiği aktif bir süreçtir; transmembran iyon gradyanlarını eski haline getirmek için ATP enerjisi gereklidir;
Aksiyon potansiyeli iletiminin mekanizması, lokal uyarımın yayılma mekanizmasından daha karmaşıktır.
Miyelinli ve miyelinsiz sinir lifleri
Miyelin lifleri. Bazı sinir lifleri embriyogenez sırasında miyelinasyona uğrar: lemositler (Schwann hücreleri) önce aksona dokunur ve sonra onu sarar (Şekil 1, A, B). Lemosit zarı akson etrafına rulo gibi sarılarak çok katmanlı bir spiral (miyelin kılıfı) oluşturur (Şekil 1, C, D). Miyelin kılıfı sürekli değildir - sinir lifinin tüm uzunluğu boyunca, birbirinden eşit mesafelerde küçük kırılmalar (Ranvier düğümleri) vardır. Engelleme alanında akson miyelin kılıfından yoksundur.
Miyelinsiz lifler. Diğer liflerin miyelinasyonu embriyonik gelişimin erken evrelerinde sona erer. Bir veya daha fazla akson lemositin içine nüfuz eder; onları tamamen veya kısmen çevreler ancak çok katmanlı bir miyelin kılıfı oluşturmaz (Şekil 1, E).
Miyelinsiz sinir lifleri boyunca uyarılma mekanizması
Dinlenme halinde, sinir lifi zarının tüm iç yüzeyi negatif yük taşır ve zarın dış tarafı pozitif yük taşır. Lipid membranın elektrik direnci yüksek olduğundan, membranın iç ve dış kısımları arasında elektrik akımı akmaz.
Bir aksiyon potansiyelinin gelişimi sırasında, zarın uyarılmış kısmında bir yük tersine dönüşü meydana gelir (Şekil 2, A). Uyarılmış ve uyarılmamış alanın sınırında bir elektrik akımı akmaya başlar (Şekil 2, B). Elektrik akımı, zarın en yakın bölümünü tahriş eder ve onu uyarılma durumuna getirir (Şekil 2, C), daha önce uyarılan alanlar ise dinlenme durumuna geri döner (Şekil 2, D). Böylece uyarılma dalgası sinir lifi zarının tüm yeni alanlarını kapsar.
Miyelinli sinir lifleri boyunca uyarılma mekanizması
Miyelinli bir sinir lifinde, miyelin kılıfıyla kaplanan membran kısımları uyarılamaz; uyarma yalnızca Ranvier düğümleri bölgesinde bulunan membran alanlarında meydana gelebilir.Ranvier düğümlerinden birinde AP'nin gelişmesiyle birlikte membran yükünün tersine çevrilmesi meydana gelir (Şekil 3, A). Membranın elektronegatif ve elektropozitif bölümleri arasında, zarın komşu bölümlerini tahriş eden bir elektrik akımı ortaya çıkar (Şekil 3, B). Bununla birlikte, yalnızca bir sonraki Ranvier düğümü alanındaki membran bölümü uyarılma durumuna girebilir (Şekil 3, B). Böylece uyarılma, membran boyunca spazmodik (tuzlayıcı) bir şekilde bir Ranvier düğümünden diğerine yayılır.
Sinir liflerinin sınıflandırılması
Sinir liflerinin çapı ve miyelinasyon derecesi farklılık gösterir. Sinir lifinin çapı ve miyelinasyon derecesi ne kadar büyük olursa, uyarılma hızı da o kadar yüksek olur. Farklı iletim hızlarına sahip lifler farklı fizyolojik işlevleri yerine getirir. Sinir lifleri, özellikleri tabloda verilen 6 tipe ayrılır. 4.1.
Tablo 4.1. Sinir lifi çeşitleri, özellikleri ve fonksiyonel amacı
Çap (μm) |
Miyelinizasyon |
İletim hızı (m/s) |
İşlevsel amaç |
|
Somatik NS'nin motor lifleri; proprioseptör duyu lifleri |
||||
Kutanöz reseptörlerin duyusal lifleri |
||||
Propriyoseptörlerin duyusal lifleri |
||||
Termoreseptörlerin, nosiseptörlerin hassas lifleri |
||||
Sempatik sinir sisteminin preganglionik lifleri |
||||
mevcut olmayan |
Sempatik sinir sisteminin postganglionik lifleri; Termoreseptörlerin, nosiseptörlerin ve bazı mekanoreseptörlerin duyusal lifleri |
Tüm grupların sinir lifleri ortak özelliklere sahiptir:
sinir lifleri neredeyse yorulmaz;
- sinir lifleri var yüksek kararsızlık yani çok yüksek frekansta bir aksiyon potansiyelini yeniden üretebilirler.

