24.06.2020
Sinteza i sekrecija, metabolizam tiroidnih hormona. Opća shema biosinteze
1. Definicija pojma "hormoni", klasifikacija i opšte biološke karakteristike hormona.
2. Klasifikacija hormona prema hemijskoj prirodi, primjeri.
3. Mehanizmi djelovanja udaljenih hormona koji prodiru u ćelije.
4. Posrednici djelovanja hormona na metabolizam - ciklični nukleotidi (cAMP, cGMP), Ca2+ joni, inozitol trifosfat, proteini receptora citosola. Reakcije sinteze i raspada cAMP.
5. Kaskadni mehanizmi aktivacije enzima kao način pojačavanja hormonskog signala. Uloga protein kinaza.
6. Hijerarhija hormonalni sistem. Princip povratne sprege u regulaciji lučenja hormona.
7. Hormoni hipotalamusa i prednje hipofize: hemijska priroda, mehanizam delovanja, ciljna tkiva i ćelije, biološki efekat.
23.1. Definicija pojma "hormoni" i njihova klasifikacija prema hemijskoj prirodi.
23.1.1. Naučite definiciju pojma: hormoni- biološki aktivni spojevi koje endokrine žlijezde izlučuju u krv ili limfu i utiču na metabolizam ćelija.
23.1.2. Zapamtite glavne karakteristike djelovanja hormona na organe i tkiva:
- hormone sintetiziraju i oslobađaju u krv specijalizirane endokrine stanice;
- hormoni imaju visoku biološku aktivnost - fiziološki učinak se manifestira kada je njihova koncentracija u krvi oko 10-6 - 10-12 mol / l;
- svaki hormon karakterizira njegova jedinstvena struktura, mjesto sinteze i funkcija; nedostatak jednog hormona ne može se nadoknaditi drugim supstancama;
- hormoni, po pravilu, utiču na organe i tkiva udaljena od mesta njihove sinteze.
23.1.3. Hormoni sprovode svoje biološko djelovanje formiranjem kompleksa sa specifičnim molekulima - receptori . Ćelije koje sadrže receptore za određeni hormon nazivaju se ciljne ćelije za ovaj hormon. Većina hormona stupa u interakciju sa receptorima koji se nalaze na plazma membrani ciljnih ćelija; drugi hormoni stupaju u interakciju sa receptorima koji se nalaze u citoplazmi i jezgru ciljnih ćelija. Imajte na umu da nedostatak i hormona i njihovih receptora može dovesti do razvoja bolesti.
23.1.4. Neki hormoni mogu biti sintetizirani od strane endokrinih stanica kao neaktivni prekursori - prohormoni . Prohormoni se mogu skladištiti u velikim količinama u posebnim sekretornim granulama i brzo aktivirati kao odgovor na odgovarajući signal.
23.1.5. Klasifikacija hormona na osnovu njihove hemijske strukture. Različite hemijske grupe hormona prikazane su u tabeli 23.1.
| Hemijska klasa | Hormon ili grupa hormona | Glavno mjesto sinteze |
|---|---|---|
| Proteini i peptidi | Liberijci statini |
Hipotalamus |
| vazopresin Oksitocin |
hipotalamus* | |
|
Tropski hormoni |
Prednja hipofiza (adenohipofiza) |
|
| Insulin Glukagon |
Gušterača (Langerhansova otočića) | |
| Parathormone | paratireoidne žlezde | |
| kalcitonin | Thyroid | |
| Derivati aminokiselina | jodotironini (tiroksin, trijodtironin) |
Thyroid |
| Kateholamini (adrenalin, norepinefrin) |
Srž nadbubrežne žlijezde, simpatički nervni sistem | |
| Steroidi | Glukokortikoidi (kortizol) |
Kora nadbubrežne žlijezde |
| Mineralokortikoidi (aldosteron) |
Kora nadbubrežne žlijezde | |
| Androgeni (testosteron) |
testisi | |
| Estrogeni (estradiol) |
jajnika | |
| Progestini (progesteron) |
jajnika |
* Mjesto lučenja ovih hormona je stražnji režanj hipofize (neurohipofiza).
Treba imati na umu da osim pravih hormona luče i one lokalni hormoni. Ove tvari sintetiziraju, po pravilu, nespecijalizirane stanice i djeluju u neposrednoj blizini mjesta proizvodnje (ne prenose se krvotokom u druge organe). Primjeri lokalnih hormona su prostaglandini, kinini, histamin, serotonin.
23.2. Hijerarhija regulatornih sistema u telu.
23.2.1. Zapamtite da postoji nekoliko nivoa regulacije homeostaze u tijelu, koji su usko povezani i funkcionišu kao jedan sistem (vidi sliku 23.1).
Slika 23.1. Hijerarhija regulatornih sistema organizma (objašnjenja u tekstu).
23.2.2. 1. Signali iz spoljašnje i unutrašnje sredine ulaze u centralni nervni sistem ( najviši nivo regulacija, vršenje kontrole unutar cijelog organizma). Ovi signali se pretvaraju u nervne impulse koji padaju na neurosekretorne ćelije hipotalamusa. Hipotalamus proizvodi:
- liberali (ili oslobađajući faktori) koji stimulišu lučenje hormona hipofize;
- statini - supstance koje inhibiraju lučenje ovih hormona.
Liberini i statini kroz sistem portalnih kapilara dospevaju do hipofize, gde se proizvode tropski hormoni . Tropski hormoni djeluju na periferna ciljna tkiva i stimuliraju (znak “+”) formiranje i lučenje hormoni perifernih endokrinih žlijezda. Hormoni perifernih žlijezda inhibiraju (znak "-") stvaranje tropskih hormona, djelujući na ćelije hipofize ili neurosekretorne ćelije hipotalamusa. Osim toga, hormoni, djelujući na metabolizam u tkivima, uzrokuju promjene u sadržaju metabolita u krvi , a oni, zauzvrat, utiču (povratnim mehanizmom) na lučenje hormona u perifernim žlijezdama (bilo direktno ili preko hipofize i hipotalamusa).
2. Nastaju hipotalamus, hipofiza i periferne žlijezde prosječan nivo regulacija homeostaze, osiguravajući kontrolu nekoliko metaboličkih puteva unutar istog organa, tkiva ili različitih organa.
Hormoni endokrinih žlijezda mogu utjecati na metabolizam:
- promjenom količine proteina enzima;
- hemijskom modifikacijom proteina enzima sa promjenom njegove aktivnosti, kao i
- promjenom brzine transporta tvari kroz biološke membrane.
3. Intracelularni regulatorni mehanizmi su najniži nivo regulacija. Signali za promjenu stanja ćelije su tvari koje se formiraju u samim stanicama ili ulaze u nju.
23.3. Mehanizmi djelovanja hormona.
29.3.1. Napominjemo da mehanizam djelovanja hormona ovisi o njegovoj kemijskoj prirodi i svojstvima - rastvorljivosti u vodi ili mastima. Prema mehanizmu djelovanja, hormoni se mogu podijeliti u dvije grupe: direktno i udaljeno djelovanje.
29.3.2. Hormoni direktnog djelovanja. U ovu grupu spadaju lipofilni (topivi u masti) hormoni - steroidi i jodotironini . Ove supstance su slabo rastvorljive u vodi i stoga formiraju kompleksna jedinjenja sa proteinima plazme u krvi. Ovi proteini uključuju i specifične transportne proteine (na primjer, transkortin, koji veže hormone kore nadbubrežne žlijezde), i nespecifične (albumine).
Hormoni direktnog djelovanja, zbog svoje lipofilnosti, mogu difundirati kroz dvostruki lipidni sloj membrane ciljnih stanica. Receptori za ove hormone nalaze se u citosolu. The emerging hormonski receptorski kompleks kreće u jezgro ćelije, gdje se vezuje za kromatin i djeluje na DNK. Kao rezultat toga, mijenja se brzina sinteze RNK na DNK šablonu (transkripcija) i brzina formiranja specifičnih enzimskih proteina na RNK šablonu (translacija). To dovodi do promjene količine enzimskih proteina u ciljnim stanicama i promjene smjera kemijskih reakcija u njima (vidi sliku 2).

Slika 23.2. Mehanizam djelovanja hormona direktnog djelovanja na ćeliju.
Kao što već znate, regulacija sinteze proteina može se provesti pomoću mehanizama indukcije i represije.
Indukcija sinteze proteina nastaje kao rezultat stimulacije sinteze odgovarajuće glasničke RNK. Istovremeno se povećava koncentracija određenog proteina-enzima u ćeliji i povećava se brzina kemijskih reakcija koje on katalizira.
Represija sinteze proteina nastaje potiskivanjem sinteze odgovarajuće RNK glasnika. Kao rezultat represije, koncentracija određenog proteina-enzima u ćeliji selektivno se smanjuje i smanjuje se brzina kemijskih reakcija koje on katalizira. Imajte na umu da isti hormon može inducirati sintezu nekih proteina i potisnuti sintezu drugih proteina. Djelovanje hormona direktnog djelovanja obično se javlja tek nakon 2 - 3 sata nakon prodiranja u ćeliju.
23.3.3. Hormoni udaljenog djelovanja. Hormoni dugog djelovanja uključuju hidrofilna (rastvorljiva u vodi) hormoni - kateholamini i hormoni proteinsko-peptidne prirode. Budući da su ove tvari netopive u lipidima, ne mogu prodrijeti kroz ćelijske membrane. Receptori za ove hormone nalaze se na vanjskoj površini plazma membrane ciljnih stanica. Udaljeni hormoni ostvaruju svoje djelovanje na ćeliju uz pomoć sekundarni posrednik, što je najčešće ciklički AMP (cAMP).
Ciklični AMP se sintetizira iz ATP-a pomoću adenilat ciklaze:

Mehanizam udaljenog djelovanja hormona prikazan je na slici 23.3.

Slika 23.3. Mehanizam djelovanja na ćelijske hormone udaljenog djelovanja.
Interakcija hormona sa svojim specifičnim receptor vodi do aktivacijaG- vjeverica stanične membrane. G-protein vezuje GTP i aktivira adenilat ciklazu.
Aktivna adenilat ciklaza pretvara ATP u cAMP, cAMP se aktivira protein kinaza.
Neaktivna protein kinaza je tetramer koji se sastoji od dvije regulatorne (R) i dvije katalitičke (C) podjedinice. Kao rezultat interakcije sa cAMP, tetramer se disocira i oslobađa se aktivni centar enzima.
Protein kinaza fosforilira enzimske proteine na račun ATP-a, ili ih aktivira ili inaktivira. Kao rezultat toga, brzina kemijskih reakcija u ciljnim stanicama se mijenja (u nekim slučajevima se povećava, u drugim smanjuje).
Inaktivacija cAMP-a nastaje uz učešće enzima fosfodiesteraze:

23.4. Hormoni hipotalamusa i hipofize.
Kao što je već spomenuto, mjesto direktne interakcije viših odjela centralne nervni sistem a endokrini sistem je hipotalamus. Ovo je mala oblast prednjeg mozga, koja se nalazi direktno iznad hipofize i povezana je sa njom sistemom krvnih sudova koji formiraju portalni sistem.
23.4.1. Hormoni hipotalamusa. Sada je poznato da neurosekretorne ćelije hipotalamusa proizvode 7 liberina(somatoliberin, kortikoliberin, tireoliberin, luliberin, foliberin, prolaktoliberin, melanoliberin) i 3 statina(somatostatin, prolaktostatin, melanostatin). Sve ove veze jesu peptidi.
Hormoni iz hipotalamusa kroz poseban portalni vaskularni sistem ulaze u prednji režanj hipofize (adenohipofiza). Liberini stimulišu, a statini potiskuju sintezu i lučenje hormona tropske hipofize. Efekat liberina i statina na ćelije hipofize je posredovan mehanizmima zavisnim od cAMP i Ca2+.
Karakteristike najčešće proučavanih liberina i statina prikazane su u tabeli 23.2.
| Faktor | Scena | Regulacija sekrecije | |
|---|---|---|---|
| Kortikoliberin | Adenohipofiza | Stimuliše lučenje adrenokortikotropnog hormona (ACTH) | Sekrecija je stimulisana stresom i potisnuta ACTH |
| Thyreoliberin | - “ - “ - | Stimuliše lučenje tireostimulirajućeg hormona (TSH) i prolaktina | Sekrecija inhibirana hormonima štitnjače |
| Somatoliberin | - “ - “ - | Stimuliše lučenje hormona rasta (STH) | Sekrecija stimulisana hipoglikemijom |
| Luliberin | - “ - “ - | Stimuliše lučenje folikulostimulirajućeg hormona (FSH) i luteinizirajućeg hormona (LH) | Kod muškaraca, lučenje je uzrokovano smanjenjem sadržaja testosterona u krvi, kod žena - smanjenjem koncentracije estrogena. Visoka koncentracija LH i FSH u krvi inhibira lučenje |
| Somatostatin | - “ - “ - | Inhibira lučenje STH i TSH | Lučenje se indukuje vježbanjem. Faktor se brzo inaktivira u tjelesnim tkivima. |
| Prolaktostatin | - “ - “ - | Inhibira lučenje prolaktina | Sekrecija je stimulisana visokom koncentracijom prolaktina i potisnuta estrogenima, testosteronom i nervnim signalima tokom sisanja. |
| Melanostatin | - “ - “ - | Inhibira lučenje MSH (hormona koji stimulira melanocite) | Sekreciju stimuliše melanotonin |
23.4.2. Hormoni adenohipofize. Adenohipofiza (prednja hipofiza) proizvodi i otpušta u krv brojne tropske hormone koji reguliraju funkciju i endokrinih i neendokrinih organa. Svi hormoni hipofize su proteini ili peptidi. Intracelularni medijator svih hormona hipofize (osim somatotropina i prolaktina) je ciklički AMP (cAMP). Karakteristike hormona prednje hipofize date su u tabeli 3.
| Hormone | ciljno tkivo | Glavni biološki efekti | Regulacija sekrecije |
|---|---|---|---|
| Adrenokortikotropni hormon (ACTH) | Kora nadbubrežne žlijezde | Stimulira sintezu i lučenje steroida u korteksu nadbubrežne žlijezde | Stimulisan kortikoliberinom |
| Tiroidni stimulirajući hormon (TSH) | Thyroid | Poboljšava sintezu i lučenje hormona štitnjače | Stimuliran tiroliberinom i potisnut hormonima štitnjače |
| Somatotropni hormon (hormon rasta, STH) | Sve tkanine | Stimulira sintezu RNK i proteina, rast tkiva, transport glukoze i aminokiselina u stanice, lipolizu | Stimulisan somatoliberinom, inhibiran somatostatinom |
| Folikul stimulirajući hormon (FSH) | Seminiferni tubuli kod muškaraca, folikuli jajnika kod žena | Povećava proizvodnju sperme kod muškaraca i formiranje folikula kod žena | Stimulisan luliberinom |
| luteinizirajući hormon (LH) | Intersticijske ćelije testisa (kod muškaraca) i jajnika (kod žena) | Izaziva lučenje estrogena, progesterona kod žena, pojačava sintezu i lučenje androgena kod muškaraca | Stimulisan luliberinom |
| Prolaktin | Mliječne žlijezde (alveolarne ćelije) | Stimuliše sintezu mlečnih proteina i razvoj mlečnih žlezda | Potisnut prolaktostatinom |
| Melanocit-stimulirajući hormon (MSH) | pigmentne ćelije | Povećava sintezu melanina u melanocitima (uzrokuje tamnjenje kože) | Potisnut melanostatinom |
23.4.3. Hormoni neurohipofize. Hormoni koje u krvotok luči stražnja hipofiza uključuju oksitocin i vazopresin. Oba hormona se sintetiziraju u hipotalamusu kao prekursorski proteini i putuju duž nervnih vlakana do stražnje hipofize.
Oksitocin - nonapeptid koji uzrokuje kontrakcije glatkih mišića materice. Koristi se u akušerstvu za stimulaciju radna aktivnost i laktacije.
vazopresin - nonapeptid koji se luči kao odgovor na povećanje osmotskog pritiska krvi. Ciljne ćelije za vazopresin su bubrežne tubularne ćelije i vaskularne glatke mišićne ćelije. Djelovanje hormona je posredovano cAMP-om. Vasopresin uzrokuje vazokonstrikciju i povećanje krvnog tlaka, a također povećava reapsorpciju vode u bubrežnim tubulima, što dovodi do smanjenja diureze.
23.4.4. Glavne vrste poremećaja hormonske funkcije hipofize i hipotalamusa. S nedostatkom somatotropnog hormona koji se javlja u djetinjstvu, razvija se patuljastost (nizak rast). Sa viškom somatotropnog hormona koji se javlja u djetinjstvu, razvija se gigantizam (nenormalno visok).
Sa viškom somatotropnog hormona koji se javlja kod odraslih (kao posljedica tumora hipofize), razvija se akromegalija - pojačan rast šaka, stopala, donje vilice, nosa.
S nedostatkom vazopresina koji je rezultat neurotropnih infekcija, razvija se traumatske ozljede mozga, tumori hipotalamusa. dijabetes insipidus. Glavni simptom ove bolesti je poliurija- naglo povećanje diureze sa smanjenom (1,001 - 1,005) relativne gustoće urina.
28.4. Hormoni pankreasa.
Imajte na umu da endokrini dio pankreasa proizvodi i oslobađa hormone inzulin i glukagon u krv.
1. Insulin. Inzulin je protein-peptidni hormon koji proizvode β-ćelije Langerhansovih otočića. Molekul insulina se sastoji od dva polipeptidna lanca (A i B) koji sadrže 21 odnosno 30 aminokiselinskih ostataka; inzulinski lanci su povezani sa dva disulfidna mosta. Inzulin se formira od prekursorskog proteina (preproinzulina) parcijalnom proteolizom (vidi sliku 4). Nakon što se signalna sekvenca cijepa, formira se proinzulin. Kao rezultat enzimske transformacije, odstranjuje se fragment polipeptidnog lanca koji sadrži oko 30 aminokiselinskih ostataka (C-peptid) i formira se inzulin.
Stimulus za lučenje inzulina je hiperglikemija - povećanje glukoze u krvi (na primjer, nakon jela). Glavne mete inzulina su ćelije jetre, mišića i masnog tkiva. Mehanizam djelovanja je udaljen.

Slika 4 Shema konverzije preproinzulina u inzulin.
insulinski receptor je kompleksni protein - glikoprotein koji se nalazi na površini ciljne ćelije. Ovaj protein se sastoji od dvije α-podjedinice i dvije β-podjedinice povezane disulfidnim mostovima. β-podjedinice sadrže nekoliko aminokiselinskih ostataka tirozina. Inzulinski receptor ima aktivnost tirozin kinaze, tj. može katalizirati prijenos ostataka fosforne kiseline iz ATP-a u OH grupu tirozina (Slika 5).

Slika 5 insulinski receptor.
U nedostatku insulina, receptor se ne pokazuje enzimska aktivnost. Kada se veže za insulin, receptor prolazi kroz autofosforilaciju, tj. β-podjedinice fosforiliraju jedna drugu. Kao rezultat toga, konformacija receptora se mijenja i on stječe sposobnost fosforilacije drugih intracelularnih proteina. Nakon toga, kompleks insulin-receptor je uronjen u citoplazmu i njegove komponente se cijepaju u lizozomima.
Formiranje kompleksa hormon-receptor povećava propusnost ćelijskih membrana za glukozu i aminokiseline. Pod dejstvom insulina u ciljnim ćelijama:
a) smanjuje se aktivnost adenilat ciklaze i povećava aktivnost fosfodiesteraze, što dovodi do smanjenja koncentracije cAMP-a;
b) brzina oksidacije glukoze se povećava, a brzina glukoneogeneze smanjuje;
c) povećava se sinteza glikogena i masti i potiskuje se njihova mobilizacija;
d) ubrzava se sinteza proteina i inhibira se njegovo raspadanje.
Sve ove promjene usmjerene su na ubrzanu upotrebu glukoze, što dovodi do smanjenja glukoze u krvi. Inaktivacija inzulina se događa uglavnom u jetri i sastoji se u razbijanju disulfidnih veza između lanaca A i B.
2. Glukagon. Glukagon je polipeptid koji sadrži 29 aminokiselinskih ostataka. Proizvode ga α-ćelije Langerhansovih otočića kao prekursorski protein (proglukagon). Djelomična proteoliza prohormona i izlučivanje glukagona u krv javlja se tijekom hipoglikemije izazvane natašte.
Ciljne ćelije za glukagon - jetra, masno tkivo, miokard. Mehanizam djelovanja je udaljen (medijator je cAMP).
Pod dejstvom glukagona u ciljnim ćelijama:
a) mobilizacija glikogena u jetri je ubrzana (vidi sliku 6) i inhibirana je njegova sinteza;
b) ubrzava se mobilizacija masti (lipoliza) u masnom tkivu i inhibira se njihova sinteza;
c) sinteza proteina je inhibirana i njihov katabolizam je pojačan;
d) ubrzana glukoneogeneza i ketogeneza u jetri.
Krajnji efekat glukagona je održavanje visokog nivoa glukoze u krvi.

Slika 6 Kaskadni mehanizam aktivacije glikogen fosforilaze pod uticajem glukagona.
3. Kršenja hormonske funkcije pankreasa. Najčešći dijabetes melitus je bolest uzrokovana kršenjem sinteze i lučenja inzulina β-ćelijama (dijabetes tipa I) ili nedostatkom receptora osjetljivih na inzulin u ciljnim stanicama (dijabetes tipa II). Dijabetes karakteriziraju sljedeći metabolički poremećaji:
a) smanjenje upotrebe glukoze u stanicama, povećanje mobilizacije glikogena i aktivacija glukoneogeneze u jetri dovode do povećanja glukoze u krvi (hiperglikemija) i njenog prevladavanja bubrežnog praga (glukozurija);
b) ubrzanje lipolize (razgradnje masti), prekomjerno stvaranje acetil-CoA, koji se koristi za sintezu sa naknadnim ulaskom u krv holesterola (hiperholesterolemija) i ketonskih tijela (hiperketonemija); ketonska tijela lako prelaze u mokraću (ketonurija);
c) smanjenje brzine sinteze proteina i povećanje katabolizma aminokiselina u tkivima dovodi do povećanja koncentracije uree i drugih dušičnih tvari u krvi (azotemija) i povećanja njihovog izlučivanja u urinu ( azoturija);
d) izlučivanje preko bubrega velike količine glukoze, ketonskih tijela i uree je praćeno povećanjem diureze (poliurija).
28.5. Hormoni medule nadbubrežne žlijezde.
Hormoni srži nadbubrežne žlijezde uključuju epinefrin i norepinefrin (kateholamini). Sintetiziraju se u hromafinskim stanicama iz tirozina (slika 7).

Slika 7 Shema za sintezu kateholamina.
Lučenje adrenalina se povećava kod stresa, fizičkog napora. Meta za kateholamine su ćelije jetre, mišićno i masno tkivo, te kardiovaskularni sistem. Mehanizam djelovanja je udaljen. Efekti se ostvaruju kroz sistem adenilat ciklaze i manifestuju se promjenama u metabolizmu ugljikohidrata. Kao i glukagon, epinefrin izaziva aktivaciju mobilizacije glikogena (vidi sliku 6) u mišićima i jetri, lipolizu u masnom tkivu. To dovodi do povećanja sadržaja glukoze, laktata i masnih kiselina u krvi. Adrenalin također pojačava srčanu aktivnost, izaziva vazokonstrikciju.
Neutralizacija adrenalina se dešava u jetri. Glavni načini neutralizacije su: metilacija (enzim - katehol-orto-metiltransferaza, COMT), oksidativna deaminacija (enzim - monoamin oksidaza, MAO) i konjugacija sa glukuronskom kiselinom. Proizvodi neutralizacije izlučuju se urinom.
Volgogradski državni medicinski univerzitetMedicinsko-biološki fakultet, III predmet
ljudska biohemija
BIOHEMIJA
STEROID
HORMONI
Slajd prezentacija predavanja
dr.sc. Valery Gennadievich Zaitsev
(Odsek za teorijsku biohemiju sa kursom kliničke biohemije, Volgogradski državni medicinski univerzitet)
© 2007, V.G. Zaitsev
Plan predavanja
UvodSteroidni hormoni - struktura, nomenklatura i
klasifikacija
Opšti pregled biosintetskih puteva steroidnih hormona
Enzimi uključeni u biosintezu
Biosintetski putevi pojedinih hormona i njihova regulacija
Steroidni hormoni u krvi
Mehanizam djelovanja/interakcija sa ciljnim stanicama
Inaktivacija i katabolizam steroidnih hormona
© 2007, V.G. Zaitsev
Karakteristike steroidnih hormona
Zajedničko porijeklo (preteča -holesterol)
Rastvorljiv u mastima, stoga lako prodire
membrane
Ne skladišti se niti čuva u endokrinom tkivu
izlučuje se odmah nakon sinteze
Regulirana sinteza, a ne oslobađanje
Enzimi za biosintezu steroidnih hormona
lokalizovan u mitohondrijama i glatki ER
Transport krvi zahtijeva posebne
proteini nosači koji se vezuju za hormone
U nekim slučajevima mogu se pretvoriti u
oblici sa promijenjenom biološkom aktivnošću
neendokrina tkiva (jetra, ciljna tkiva)
© 2007, V.G. Zaitsev
Kostur steroidnih hormona
1,2-Ciklopentanoperhidrofenantren4 ugljovodonična prstena
(3 šestočlana i 1
petočlani)
Alternativni položaji
označeno strelicama
Moguće zamjene:
metil-, hidroksi-, okso-,
karboksil-, acetil-,
hidroksiacetil-,
karboksialkil i drugi.
© 2007, V.G. Zaitsev
Metaboličko porijeklo
Svi steroidni hormoni– lipofilna
niske molekularne težine
veze, zajednički
prethodnik
koji je
holesterol
Izvori holesterola
ljudsko tijelo:
hrana i biosinteza
(uglavnom u
hepatociti)
© 2007, V.G. Zaitsev
Mjesto steroidnih hormona u metabolizmu holesterola
KOLESTEROLŽučne kiseline
Progesteron
Glukokortikoidi
Mineralokortikoidi
vitamin D
Androgeni
Estrogeni
© 2007, V.G. Zaitsev
Stereohemija steroida
Za B/C komunikacijuživotinjski steroidi
poznata samo cis konformacija
A/B i C/D veze
može biti i cis i
trans-većina
steroid
ljudski hormoni
imaju konformaciju
trans-trans-trans
(5α-steroidi)
5α-steroidi
5β-steroidi
© 2007, V.G. Zaitsev
Regulatorni steroidi
1.2.
"Pravi" steroidni hormoni:
sintetišu se uglavnom u unutrašnjim žlezdama
sekreta
endokrini efekti
Neurosteroidi (Baulieu E.E., 1991; Biol. Cell. 71:3-10)
sintetiziraju ćelije centralnog nervnog sistema
(CNS)
autokrini i parakrini efekti
© 2007, V.G. Zaitsev
Neophodni koraci razmjene
Sinteza steroidnih hormona direktno iz holesterolaili iz međuproizvoda
Lučenje steroidnih hormona u krv / transport do ciljeva
akcije
Periferni metabolizam (konverzija primarnog steroida
hormone u metabolite s drugom biološkom aktivnošću,
javlja se u jetri i ciljnim ćelijama)
Upijanje od strane ciljnih ćelija
Inaktivacija i katabolizam steroidnih hormona / izlučivanje
proizvodi raspadanja
© 2007, V.G. Zaitsev
10. Klasifikacije steroidnih hormona
Po mjestu školovanjaPo biološkim funkcijama (koji sistemi
utjecati)
Po vrstama biološke aktivnosti
Biohemijskom aktivnošću
Po tipu ciljne ćelije
Po hemijskoj strukturi
Po spolu (univerzalno/muško/žensko)
© 2007, V.G. Zaitsev
11. Mjesta obrazovanja
ADRENAL (kortikosteroidi)glukokortikoidi i mineralokortikoidi, dijelom -
progesteron i neki androgeni)
TESTISI (muški polni hormoni - androgeni)
JAJNICI (ženski polni hormoni - progestini i
estrogen)
FETOPLANTENTNO ENDOKRINO TKIVO
(progesteron od 6-8 nedelje trudnoće, kao i
estrogeni - iz dehidroepiandrosteron sulfata)
© 2007, V.G. Zaitsev
12. Klase steroidnih hormona
GLUKOKORTIKOIDI (glavni predstavnik -kortizol)
MINERALOKORTIKOIDI (najpoznatiji i proučavani
aldosteron)
ANDROGENI (npr. testosteron)
PROGESTINI, ili PROGESTOGENI (progesteron)
ESTROGENI (najznačajniji su estradiol i estron)
© 2007, V.G. Zaitsev
13. Opća shema biosinteze
© 2007, V.G. Zaitsev14. Uobičajeni metabolički prekursor
Pregnenolon (C21 steroid)Nastaje u prvoj fazi sinteze
SVI steroidni hormoni
Reakcija cijepanja bočnog lanca
kolesterol katalizira specijalna
ovisni o citokromu P450
enzim - R450scc (također
naziva se 20,22-desmolaza ili
20,22-lyase)
Ključni korak u sintezi steroida
hormoni
Regulira adrenokortikotropno
hormona (ACTH) u nadbubrežnim žlijezdama i
luteinizirajući hormon (LH) u
gonade
© 2007, V.G. Zaitsev
15. Androgeni steroidi
Testosteron17β-hidroksiandrost-4-en-3-on
Androstenedione
Androst-4-en-3,17-dion
5α-dihidrotestosteron
17β-hidroksi-5β-androstan-3-on
© 2007, V.G. Zaitsev
16. Androgeni steroidi
Mjesta sintezetestisi
Kora nadbubrežne žlijezde
Androgena aktivnost
Rast i razvoj muških reproduktivnih organa
Uključen je u određivanje spola fetusa
Utječu na spolno specifično ponašanje
Odrediti ispoljavanje sekundarnih polnih karakteristika
Stimulansi i regulatori spermatogeneze
Anaboličko djelovanje
Razvoj mišićne mase
Razvoj skeleta i vezivnog tkiva
Razvoj kose
Izazivaju preokret kataboličkih procesa,
što dovodi do smanjenja mase određenih vrsta tkiva
Stimulacija sinteze proteina, suzbijanje njegovog razlaganja
© 2007, V.G. Zaitsev
17. Anabolički steroidi
© 2007, V.G. Zaitsev18. Estrogeni steroidi
EstronEstradiol
3-hidroksiestr-1,3,5-trien-17-on
estr-1,3,5-trien-3,17β-diol
Estriol
Estr-1,3,5-trien-3,16α,17β-triol
© 2007, V.G. Zaitsev
19. Estrogeni steroidi
Mjesta sintezejajnika
Placenta
U malim količinama - nadbubrežne žlijezde, hipotalamus,
adenohipofiza, testis
Fiziološka aktivnost prirodnih estrogena
Regulacija reprodukcije
Razvoj ženskih reproduktivnih organa
Regulacija ovulacije
Priprema ženskog tijela za trudnoću
regulisanje faza trudnoće
Regulacija metabolizma koštanog tkiva (rast)
Regulacija prirode tjelesne masti
© 2007, V.G. Zaitsev
20. Sintetički estrogeni
Jači od prirodnih estrogena, potiskujuovulacija
Uključeno u oralne kontraceptive
© 2007, V.G. Zaitsev
21. Progestini
Mjesta sintezeŽuto tijelo jajnika
Placenta
testisi
Kora nadbubrežne žlijezde
Fiziološka aktivnost
prirodni estrogeni
Očuvanje i održavanje
trudnoća
potiskivanje sazrevanja
folikula i ovulacije
Prevencija spontanih
kontrakcije materice
Razvoj grudi
Progesteron
preg-4-en-3,20-dion
© 2007, V.G. Zaitsev
22. Mineralokortikoidi
Mjesta sintezeKora nadbubrežne žlijezde (zona glomerulosa)
Fiziološka aktivnost
Regulacija nivoa i ravnoteže elektrolita (pojačati
reapsorpcija natrijuma i izlučivanje kalija)
Regulacija razmjene vode
Porast krvnog pritiska
aldehidni oblik
hemiacetalni oblik
Aldosteron
11β,21-dihidroksipregn-4-en-3,18,20-trion
© 2007, V.G. Zaitsev
23. Glukokortikoidi
Mjesta sintezeKora nadbubrežne žlijezde (zona fasciculata)
Fiziološka aktivnost
Regulacija metabolizma ugljikohidrata (glukoneogeneza), proteina
(proteoliza), masti (lipoliza), kalcijum
suzbijanje aktivnosti imunološki sistem, regulacija,
upalne i alergijske reakcije
Neki od hormona stresa
Uključen u formiranje pamćenja, učenja,
raspoloženja, cirkadijalni ritmovi
© 2007, V.G. Zaitsev
24. Dnevni ritam lučenja kortizola
© 2007, V.G. Zaitsev25. Regulatori sinteze steroidnih hormona
1.luteinizirajući hormon (LH)
progesteron i testosteron
2.
Adrenokortikotropni hormon (ACTH)
kortizol
3.
Folikul stimulirajući hormon (FSH)
estrogeni
4.
Angiotenzini II i III
aldosteron
© 2007, V.G. Zaitsev
26. Steroidogeni enzimi
nalazi se u mitohondrijama i glatkom ER1.
2.
3.
4.
desmolaze (lyases)
P450scc uklanja dio bočnog lanca holesterola. Reakcija
zahtijeva citokrom P450, O2, NADPH. Enzim
mitohondrijski, zajedno sa transportom elektrona
sistem
Hidroksilaze
Zahtijeva citokrom P450, O2, NADPH i može biti
nalaze u mitohondrijima i ER
Dehidrogenaze hidroksiliranih steroida
(oksidoreduktaza)
Može biti citosolna ili mikrozomalna. Reakcije
reverzibilan, smjer ovisi o omjeru
NAD(P)/NAD(P)H
Aromataza
Transformiše A prsten u aromatični prsten. Enzim vezan za citokrom P450 vezan na membranu
© 2007, V.G. Zaitsev
27. Steroidogeni enzimi
Trivijalno ime"stari"
oznaka
"Novo"
oznaka
Desmolase
P450scc
CYP11A1
3β-hidroksisteroid dehidrogenaza
3β-HSD
3β-HSD
17α-Hydroxylase / 17,20 lyase
P450C17
CYP17
21-Hydroxylase
P450C21
CYP21A2
11β-Hydroxylase
P450C11
CYP11B1
Aldosteron synthase
P450C11AS
CYP11B2
Aromataza
P450aro
CYP19
© 2007, V.G. Zaitsev
28. Steroidogeni enzimi
© 2007, V.G. Zaitsev29. Sinteza steroida u nadbubrežnim žlijezdama
* DHEA-S - dehidroepiandrosteron sulfat© 2007, V.G. Zaitsev
30. Regulacija sinteze steroida u nadbubrežnim žlijezdama
zona fasciculata + zona reticularisadrenokortikotropni hormon (ACTH) + kortikotropinliberin + kortizol (negativna povratna informacija)
cAMP zavisna regulacija
zona glomerulosa
angiotenzini II i III stimulišu P450scc
regulacija nivoa intracelularnog Ca2+ putem
mehanizam ovisan o protein kinazi C
Kalijum u plazmi može regulisati sintezu
mineralokortikoidi direktno, kroz djelovanje
naponski zavisni Ca2+ kanali
promjena nivoa kalijuma u plazmi za samo 0,1 mM
uzrokuje skoro dvostruku promjenu sekrecije
aldosteron
© 2007, V.G. Zaitsev
31. Sinteza polnih hormona
© 2007, V.G. Zaitsev32. Regulacija sinteze androgena
MENLeydig ćelije
LH stimulira proizvodnju testosterona pomoću cAMP-ovisnog mehanizma
ne mogu sintetizirati dihidrotestosteron
Sertolijeve ćelije
Proizvodnja testosterona je stimulirana FSH preko cAMP-ovisnog mehanizma
mogu koristiti endogene i egzogene (iz stanica
Leydig) testosteron za sintezu dihidrotestosterona
ŽENE
Tekalne ćelije jajnika
proizvodnju androstendiona i testosterona
stimuliran LH mehanizmom zavisnim od cAMP
© 2007, V.G. Zaitsev
33. Regulacija sinteze androgena
* StAR - Steroidogenic Acute Regulatory Protein© 2007, V.G. Zaitsev
34. Androgeni antagonisti
CH3OHO
CH3
CH
CH3
N
H
CH3
CH3
CH3
CH3
N
O
O
N
HH
Finesterid
Danazol
(ćelavost)
(endometrioza)
O
O
O
CH3
O
HN
S
HN
HO
CH3
CH3
F
CF3
CF3
CN
NO2
Bikalutamid
Flutamid
(rak prostate)
(rak prostate)
© 2007, V.G. Zaitsev
35. Aromataza u sintezi estrogena
Aromataza je prisutna u tekalnim i granuloza ćelijamajajnika
U tekalnim stanicama, sinteza estrogena (lučenje u krv)
stimulira LH kroz aktivaciju sinteze androgena
U granularnoj sintezi estrogena (iz tekalnih androgena
ćelije, izlučivanje u folikularnu tečnost)
stimuliran FSH kroz povećanu aktivnost
aromataza. Sazrijevanje ćelija granuloze povećava njihovu
preosjetljivost na LH
© 2007, V.G. Zaitsev
36. Aromataza u sintezi estrogena
OCYP19
O
CYP19
O
HO
H
HO
HO
O2, NADPH
O
O2, NADPH
O
O
androstenedione
19,19-dihidroksiandrostendion
19-hidroksiandrostendion
O
-H2O
+3
Fe
O
-HCOOH
HO
O
CYP19
O
O2, NADPH
H2O
O
O
HO
O
estron
O
peroksi enzim
srednji
19-oksoandrostendion
© 2007, V.G. Zaitsev
37. Aromataza u sintezi estrogena
OH
O
androstenedione
Oh
17-HSD
Oh
aromataza
HO
+H2O
+
HCOOH
O
estradiol
testosteron
© 2007, V.G. Zaitsev
44. Receptori za steroidne hormone
(intranuklearni)
dimerizacija
(intracelularno)
transkripcija
prevod
Intracelularni efekti
proteini
ekstracelularni efekti
© 2007, V.G. Zaitsev 45. Steroidni hormonski receptori
S
R
hsp hsp
R
S
+
hsp hsp
R
S
R
+ili-
S
HRE
Ciljni gen
© 2007, V.G. Zaitsev 46. Receptori za steroidne hormone
Citoplazma
Ligand
Nukleus
ćelijski specifičan
Odgovor
SR
SR SR
proteina
SR
TF TF
SR SR
HRE
mRNA
© 2007, V.G. Zaitsev 47. Regulacija funkcije steroidnih hormona
Koncentracija hormona
Fosforilacija/defosforilacija
Pri niskim koncentracijama steroida dolazi do fosforilacije
obično slaba
Fosforilirani serin i treonin
Enzim: protein kinaze aktivirane mitogenom
(MAPK-ovi)
Vezivanje steroida može povećati stepen
fosforilacija
Fosforilacija povećava afinitet receptora za
DNK, transkripciona aktivnost i stabilnost
hormon-receptor-DNK kompleks
© 2007, V.G. Zaitsev 48. Receptori za steroidne hormone
© 2007, V.G. Zaitsev 49. Receptori za steroidne hormone
E
ER
E
ER
element odgovora na estrogen
E
ER
Fos
juna
AP-1 element (ili Sp-1)
© 2007, V.G. Zaitsev 50.
Steroidni receptori Receptori klase II
GR Glucocorticoid
PR Progesteron
AR Android
ER Estrogen
SR
SR
Palindrom HREs
Orphan Receptors
VDR, PPAR
TR, FXR
RXR, LXR
RAR, PXR
NR RXR
Direktno ponavljanje HRE
NGFI-B
SF-I
ERR
ReVERB
NR
HRE na pola lokacije
AAA-ACGGTCA NBRE
AGAACA-N3-TGTTCT GRE/PRE ACGGTCA-N1-5-AGGTCA
TCA-AGGTCA SFRE
AGGTCA-N3-TGACCT ERE 51. Koaktivatori estrogenskih receptora
CBP
pCAF
acetilacija histona
SRC SRC
ER
RGGTCA
ER
ACTGGR
TFII-B
TBP
transkripcija
RNA
Paul
© 2007, V.G. Zaitsev 52. Sinergija hormona
© 2007, V.G. Zaitsev 53. Inaktivacija steroidnih hormona
© 2007, V.G. Zaitsev 54. Bolesti povezane sa poremećajima metabolizma steroida
HIRSUTIZAM (višak proizvodnje
dehidroepiandrosteron, defekt u jednom od 3
biosintetski enzimi)
ADDISONOVA BOLEST (hipokorticizam)
CUSHINGOV SINDROM (hiperkortizolizam - tumori
nadbubrežna ili hipofizna, jatrogena)
HIPERKORTIZAM bez Cushingovog sindroma
SINDROM NEOSJETLJIVOSTI NA ANDROGE
(feminizacija testisa)
© 2007, V.G. Zaitsev
Normalna fiziologija Marina Gennadievna Dragoy
27. Sinteza, lučenje i izlučivanje hormona iz organizma
Biosinteza hormona je lanac biohemijskih reakcija koje formiraju strukturu hormonskog molekula. Ove reakcije se odvijaju spontano i genetski su fiksirane u odgovarajućim endokrinim ćelijama.
Genetska kontrola se provodi ili na nivou formiranja mRNA (messenger RNA) samog hormona ili njegovih prekursora, ili na nivou formiranja mRNA proteina enzima koji kontrolišu različite faze formiranja hormona.
Ovisno o prirodi hormona koji se sintetiše, postoje dvije vrste genetske kontrole hormonske biogeneze:
1) direktna, biosintezna shema: "geni - mRNA - prohormoni - hormoni";
2) posredovano, shema: "geni - (mRNA) - enzimi - hormon".
Lučenje hormona - proces oslobađanja hormona iz endokrinih ćelija u međućelijske praznine sa njihovim daljim ulaskom u krv, limfu. Lučenje hormona je striktno specifično za svaku endokrinu žlijezdu.
Proces sekrecije se odvija i u mirovanju i pod uslovima stimulacije.
Lučenje hormona se odvija impulsivno, u odvojenim diskretnim porcijama. Impulzivna priroda hormonske sekrecije objašnjava se cikličnom prirodom procesa biosinteze, taloženja i transporta hormona.
Sekrecija i biosinteza hormona usko su međusobno povezane. Ovaj odnos zavisi od hemijske prirode hormona i karakteristika mehanizma sekrecije.
Postoje tri mehanizma lučenja:
1) oslobađanje iz ćelijskih sekretornih granula (lučenje kateholamina i proteinsko-peptidnih hormona);
2) oslobađanje iz oblika vezanog za proteine (lučenje tropskih hormona);
3) relativno slobodna difuzija kroz ćelijske membrane (lučenje steroida).
Stepen povezanosti između sinteze i lučenja hormona raste od prve do treće vrste.
Hormoni, ulazeći u krv, prenose se do organa i tkiva. Vezani za proteine plazme i oblikovani elementi hormon se akumulira u krvotoku, privremeno se isključuje iz kruga biološkog djelovanja i metaboličkih transformacija. Neaktivni hormon se lako aktivira i dobija pristup ćelijama i tkivima.
Paralelno, postoje dva procesa: implementacija hormonskog efekta i metabolička inaktivacija.
U procesu metabolizma hormoni se mijenjaju funkcionalno i strukturno. Velika većina hormona se metabolizira, a samo mali dio (0,5-10%) se izlučuje nepromijenjen. Metabolička inaktivacija se najintenzivnije javlja u jetri, tanko crijevo i bubrezi. Proizvodi hormonskog metabolizma se aktivno izlučuju u urinu i žuči, a žučne komponente se konačno izlučuju fecesom kroz crijeva.
autor Marina Gennadievna Dragoj Iz knjige Homeopatija. Dio II. Praktične preporuke na izbor lekova od Gerharda Kelera Iz knjige Osnovi intenzivne rehabilitacije. povreda kičme i kičmena moždina autor Vladimir Aleksandrovič Kačesov Iz knjige Normalna fiziologija autor Iz knjige Normalna fiziologija autor Nikolaj Aleksandrovič Agadžanjan Iz knjige Atlas: ljudska anatomija i fiziologija. Završeno praktični vodič autor Elena Yurievna Zigalova Iz knjige Filozofski kamen homeopatije autor Natalija Konstantinovna Simeonova Iz knjige Iscjeljujuće sile. Knjiga 1. Čišćenje organizma i pravilnu ishranu. Biosinteza i bioenergetika autor Genadij Petrovič Malahov Iz knjige Tajne iscjelitelja Istoka autor Viktor Fedorovič Vostokov Iz knjige Thalasso and Relaxation autor Irina Krasotkina autor Boris Vasiljevič Bolotov Iz knjige Bolotovljevi recepti za svaki dan. Kalendar za 2013 autor Boris Vasiljevič Bolotov autor Galina Ivanovna ujak Iz knjige Kako uravnotežiti svoje hormone štitne žlijezde, nadbubrežne žlijezde, pankreas autor Galina Ivanovna ujak Iz knjige Ljekoviti čajevi autor Mikhail Ingerleib Iz knjige Minimum masnoće, maksimum mišića! od Maxa LisaPoglavlje 16
Koncept hormona. Osnovni principi regulacije metabolizma
Jedna od jedinstvenih osobina živih organizama je njihova sposobnost da održavaju postojanost homeostaze (stalnost mnogih svojstava tijela u stalnim uvjetima okoline) koristeći mehanizme samoregulacije, u čijoj koordinaciji jedno od glavnih mjesta pripada hormonima. . Hormoni su biološki aktivne tvari organske prirode koje se proizvode u stanicama endokrinih žlijezda i imaju regulatorni učinak na metabolizam.
Kao rezultat djelovanja mehanizama samoregulacije, odnosno neurohormonalnih mehanizama, u živoj ćeliji se međusobno usklađuju brzine svih hemijskih reakcija i fizičko-hemijskih procesa, koordinacija funkcija svih organa i adekvatan odgovor tijela na promjene u vanjskoj sredini su osigurane. U regulaciji metaboličkih procesa hormoni zauzimaju međupoziciju između nervnog sistema i delovanja enzima, tj. regulacija metabolizma se ostvaruje promjenom brzine enzimskih reakcija. Hormoni izazivaju ili vrlo brzu reakciju, ili obrnuto, sporu reakciju povezanu sa ponovnom sintezom potrebnog enzima. Dakle, poremećaji u sintezi i razgradnji hormona, uzrokovani, na primjer, bolestima endokrinih žlijezda, dovode do promjene u normalnoj sintezi enzima i, posljedično, do metaboličkih i energetskih poremećaja.
U mehanizmima samoregulacije mogu se razlikovati tri nivoa.
Prvi nivo - intracelularni mehanizmi regulacije. Različiti metaboliti služe kao signali za promjenu stanja ćelije. Oni mogu:
- mijenjaju aktivnost enzima tako što ih inhibiraju ili aktiviraju;
- mijenjaju količinu enzima regulacijom njihove sinteze i raspadanja;
- promijeniti brzinu transmembranskog iskrivljenja tvari. Međuorganska koordinacija ovog nivoa regulacije obezbeđuje se prenosom signala na dva načina: kroz krv uz pomoć hormona (endokrini sistem) i preko nervnog sistema.
Drugi nivo regulacije - endokrini sistem. Hormoni se oslobađaju u krv kao odgovor na određeni podražaj, koji može biti nervni impuls ili promjena koncentracije nekog metabolita u krvi koja teče kroz endokrinu žlijezdu (na primjer, smanjenje koncentracije glukoze). Hormon se transportuje krvlju i, dostižući ciljne ćelije, modifikuje njihov metabolizam kroz unutarćelijske mehanizme. U tom slučaju dolazi do promjene metabolizma i eliminira se podražaj koji je izazvao oslobađanje hormona. Hormon koji je izvršio svoju funkciju uništavaju posebni enzimi.
Treći nivo regulacije je nervni sistem sa receptorima za signale kako iz spoljašnje, tako i unutrašnje sredine. Signali se transformišu u nervni impuls, koji u sinapsi sa efektorskom ćelijom izaziva oslobađanje posrednika – hemijskog signala. Posrednik kroz intracelularne mehanizme regulacije izaziva promjenu metabolizma. Efektorske ćelije mogu biti i endokrine ćelije koje na nervni impuls reaguju sintezom i oslobađanjem hormona.
Sva tri nivoa regulacije su međusobno usko povezana i deluju kao jedan neuro-hormonski ili neuro-humoralni sistem regulacije (Sl. 43).
Tok informacija o stanju vanjskog i unutrašnjeg okruženja tijela ulazi u nervni sistem, gdje se obrađuje, a kao odgovor se šalju regulatorni signali do perifernih organa i tkiva. Pod direktnom kontrolom nervnog sistema su medula nadbubrežne žlezde i hipotalamus. Nervni impulsi koji dolaze iz različitih dijelova mozga utiču na lučenje neuropeptida od strane hipotalamičkih stanica - liberina i statina, koji reguliraju oslobađanje tropskih hormona iz hipofize. Liberini stimulišu sintezu i oslobađanje trostrukih hormona, dok je statini inhibiraju. Trostruki hormoni hipofize utiču na lučenje hormona u perifernim žlezdama. Stvaranje i lučenje hormona od strane perifernih žlijezda odvija se kontinuirano. To je neophodno za održavanje željenog nivoa u krvi, jer se brzo inaktiviraju i izlučuju iz organizma.
Rice. 43. Šema neurohormonske regulacije (pune strelice označavaju sintezu hormona, a tačkaste strelice pokazuju uticaj hormona na ciljne organe)
Koncentracija hormona u krvi je niska: oko 10 -6 - 10 - 11 mol/l. Poluvrijeme je obično nekoliko minuta, za neke je desetine minuta, vrlo rijetko sati. Potreban nivo hormona u krvi održava se mehanizmom samoregulacije po plus ili minus principu međuhormonskih odnosa. Tropski hormoni potiču stvaranje i lučenje hormona od strane perifernih žlijezda (znak "+"), a potonje mehanizmom negativne povratne sprege inhibiraju (znak "-") stvaranje tropskih hormona, djelujući preko hipofize. ćelije (kratka povratna sprega) ili neurosekretorne ćelije hipotalamusa (duga povratna sprega), sl.44. U potonjem slučaju inhibira se lučenje liberina u hipotalamusu.

Osim toga, postoji metabolitsko-hormonska povratna sprega: hormon, djelujući na metabolizam u tkivima, uzrokuje promjenu sadržaja bilo kojeg metabolita u krvi, a koji mehanizmom povratne sprege utiče na lučenje hormona u perifernom žlezde ili direktno (intracelularni mehanizam), ili preko hipofize i hipotalamusa (vidi sliku 44). Takvi metaboliti su glukoza (pokazatelj stanja metabolizma ugljikohidrata), aminokiseline (indikator stanja metabolizma proteina), nukleotidi i nukleozidi (indikatori stanja metabolizma nukleinskih i proteina), masne kiseline, kolesterol (indikatori za stanje metabolizma lipida); H 2 O, Ca 2+, Na+, K+, CI¯ i neki drugi joni (indikatori stanja ravnoteže vode i soli).
Klasifikacija hormona
Hormoni imaju sljedeće zajedničke biološke karakteristike:
1) dismorfno dejstvo, odnosno regulišu metabolizam i funkcije efektorskih ćelija na daljinu;
2) stroga specifičnost biološkog delovanja, odnosno da se jedan hormon ne može u potpunosti zameniti drugim;
3) visoka biološka aktivnost - vrlo male količine, ponekad i desetak mikrograma, dovoljne su da se organizam održi u životu.
Hormoni se klasifikuju prema:
1) hemijske prirode;
2) mehanizam prenosa signala u ćeliju - cilj;
3) biološke funkcije.
Sve vrste klasifikacije su nesavršene i donekle proizvoljne, posebno klasifikacija po funkciji, jer su mnogi hormoni polifunkcionalni.
Po hemijskoj strukturi hormoni se dijele na sljedeći način:
1) protein-peptid (hormoni hipotalamusa, hipofize, pankreasa i paratireoidnih žlezda, kalciotonin štitne žlezde);
2) derivati aminokiselina (adrenalin je derivat fenilanina i tirozina);
3) steroidi (spolni hormoni - androgeni, estrogeni i gestageni, kortikosteroidi).
Prema biološkim funkcijama Hormoni se dijele u sljedeće grupe:
1) regulisanje metabolizma ugljenih hidrata, masti, aminokiselina - insulina, glukagona, adrenalina, glukokortikosteroida (kortizola);
2) regulisanje metabolizma vode i soli - mineralokortikosteroidi (aldosteron), antidiuretski hormon (vazopresin);
3) regulisanje razmene kalcijuma i fosfata - paratiroidnog hormona, kalcitonina, kalcitriola;
4) regulisanje metabolizma povezanog sa reproduktivnom funkcijom (spolni hormoni) - estradiol, progesteron, testosteron.
5) regulatorne funkcije endokrinih žlijezda (trostruki hormoni) - kortikotropin, tirotropin, gonadotropin.
Ova klasifikacija ne uključuje somatotropin, tiroksin i neke druge hormone koji imaju polifunkcionalno djelovanje.
Osim toga, pored hormona koji se oslobađaju u krv i djeluju na organe udaljene od mjesta sinteze hormona, postoje i lokalni hormoni koji regulišu metabolizam u onim organima u kojima se formiraju. Tu spadaju hormoni gastrointestinalnog trakta, hormoni mastocita vezivnog tkiva (heparin, histamin), hormoni koje luče ćelije bubrega, sjemenih vezikula i drugih organa (prostaglandini) itd.
Slične informacije.
Struktura modula | Teme |
Modularna jedinica 1 | 11.1. Uloga hormona u regulaciji metabolizma 11.2. Mehanizmi prijenosa hormonskih signala do stanica 11.3. Struktura i sinteza hormona 11.4. Regulacija razmjene glavnih energetskih nositelja uz normalan ritam ishrane 11.5. Promjene u metabolizmu tijekom hipo- i hipersekrecije hormona |
Modularna jedinica 2 | 11.6. Promjene hormonalni status i metabolizam natašte 11.7. Promjene u hormonskom statusu i metabolizmu kod dijabetes melitusa |
Modularna jedinica 3 | 11.8. Regulacija metabolizma vode i soli 11.9. Regulacija metabolizma kalcijuma i fosfata. Struktura, sinteza i mehanizam djelovanja paratiroidnog hormona, kalcitriola i kalcitonina |
Modularna jedinica 1 ULOGA HORMONA U REGULACIJI METABOLIZMA. REGULACIJA METABOLIZMA UGLJENIH HIDRATA, LIPIDA, AMINOKISELINA NORMALNIM RITMOM ISHRANA
Ciljevi učenja Biti u stanju:
1. Primijeniti znanje o molekularnim mehanizmima regulacije metabolizma i tjelesnih funkcija da biste razumjeli biohemijske osnove homeostaze i adaptacije.
2. Iskoristiti znanje o mehanizmima djelovanja hormona (insulin i kontranzularni hormoni: glukagon, kortizol, adrenalin, somatotropin, jodotironin) za karakterizaciju promjena u energetskom metabolizmu pri promjeni perioda probave i postapsorptivnog stanja.
3. Analizirati promjene u metabolizmu tokom hipo- i hiperprodukcije kortizola i hormona rasta, Itsenko-Cushingove bolesti i sindroma (akromegalija), kao i hiper- i hipofunkcije štitne žlijezde (difuzna toksična struma, endemska struma).
znati:
1. Savremena nomenklatura i klasifikacija hormona.
2. Glavne faze prenosa hormonskih signala u ćeliju.
3. Faze sinteze i sekrecije inzulina i glavnih kontrainsularnih hormona.
4. Mehanizmi za održavanje koncentracije glavnih nosilaca energije u krvi
telad sa normalnim ritmom ishrane.
Tema 11.1. ULOGA HOMONA U REGULACIJI METABOLIZMA
1. Za normalno funkcioniranje višećelijskog organizma neophodan je odnos između pojedinih stanica, tkiva i organa. Ovaj odnos se ostvaruje:
nervni sistem(centralni i periferni) putem nervnih impulsa i neurotransmitera;
endokrini sistem preko endokrinih žlijezda i hormoni, koje sintetiziraju specijalizirane stanice ovih žlijezda, oslobađaju se u krv i transportuju do različitih organa i tkiva;
parakrina i autokrini sistema putem različitih jedinjenja koja se luče u međućelijski prostor i stupaju u interakciju sa receptorima bilo obližnjih ćelija ili iste ćelije (prostaglandini, hormoni gastrointestinalnog trakta, histamin, itd.);
imuni sistem preko specifičnih proteina (citokina, antitijela).
2. Endokrini sistem osigurava regulaciju i integraciju metabolizma u različitim tkivima kao odgovor na promjene u uvjetima vanjskog i unutrašnjeg okruženja. Hormoni funkcioniraju kao kemijski glasnici koji prenose informacije o ovim promjenama u različite organe i tkiva. Odgovor ćelije na djelovanje hormona određen je kako hemijskom strukturom hormona tako i vrstom ćelije na koju je njegovo djelovanje usmjereno. Hormoni su prisutni u krvi u vrlo niskim koncentracijama i njihovo djelovanje je obično kratkotrajno.
To je zbog, prvo, regulacije njihove sinteze i sekrecije i, drugo, visoke stope inaktivacije cirkulirajućih hormona. Glavne veze između nervnog i endokrinog sistema regulacije se odvijaju uz pomoć posebnih dijelova mozga - hipotalamusa i hipofize. Sistem neurohumoralne regulacije ima svoju hijerarhiju, čiji je vrh CNS i strogi slijed procesa.
3. Hijerarhija regulatornih sistema. Sistemi za regulaciju metabolizma i tjelesnih funkcija čine tri hijerarhijska nivoa (slika 11.1).
Prvi nivo- centralnog nervnog sistema. Nervne ćelije primaju signale iz spoljašnje i unutrašnje sredine, pretvaraju ih u oblik nervnog impulsa, koji u sinapsi izaziva oslobađanje medijatora. Medijatori uzrokuju metaboličke promjene u efektorskim stanicama kroz intracelularne regulatorne mehanizme.
Drugi nivo- endokrini sistem- obuhvata hipotalamus, hipofizu, periferne endokrine žlijezde, kao i specijalizirane ćelije nekih organa i tkiva (gastrointestinalni trakt, adipociti), koje sintetiziraju hormone i ispuštaju ih u krv pod djelovanjem odgovarajućeg stimulusa.
Treći nivo- intracelularno- predstavljaju promjene u metabolizmu unutar ćelije ili određenog metaboličkog puta koje su rezultat:
Promjene aktivnost enzimi aktivacijom ili inhibicijom;
Promjene količine enzimi mehanizmom indukcije ili potiskivanja sinteze proteina ili promjena u brzini njihove degradacije;
Promjene brzina transporta tvari kroz ćelijske membrane. Sinteza i lučenje hormona stimulisan spoljašnjim i unutrašnjim
signale CNS-u. Ovi signali preko nervnih veza ulaze u hipotalamus, gdje stimuliraju sintezu peptidnih hormona (tzv. oslobađajućih hormona) - liberina i statina. Liberijci i statini transportuju se u prednju hipofizu, gde stimulišu ili inhibiraju sintezu tropskih hormona. Tropski hormoni hipofize potiču sintezu i lučenje hormona iz perifernih endokrinih žlijezda, koji ulaze u opću cirkulaciju. Neki hormoni hipotalamusa se pohranjuju u stražnjoj hipofizi, odakle se izlučuju u krv (vazopresin, oksitocin).
 Promjena koncentracije metabolita u ciljnim stanicama mehanizmom negativne povratne sprege potiskuje sintezu hormona, djelujući ili na endokrine žlijezde ili na hipotalamus; sintezu i lučenje tropskih hormona potiskuju hormoni perifernih žlijezda.
Promjena koncentracije metabolita u ciljnim stanicama mehanizmom negativne povratne sprege potiskuje sintezu hormona, djelujući ili na endokrine žlijezde ili na hipotalamus; sintezu i lučenje tropskih hormona potiskuju hormoni perifernih žlijezda.
TEMA 11.2. MEHANIZMI PRENOSA HORMONSKIH SIGNALA U ĆELIJE
Biološko djelovanje hormona manifestuje se kroz njihovu interakciju sa ćelijama koje imaju receptore za ovaj hormon (ciljne ćelije). Da bi bio biološki aktivan, vezivanje hormona za receptor mora rezultirati hemijskim signalom unutar ćelije koji izaziva specifičan biološki odgovor, kao što je promjena brzine sinteze enzima i drugih proteina ili promjena njihove aktivnosti ( vidi Modul 4). Meta za hormon mogu poslužiti kao ćelije jednog ili više tkiva. Utječući na ciljnu ćeliju, hormon izaziva specifičan odgovor, čija manifestacija ovisi o tome koji su metabolički putevi aktivirani ili inhibirani u ovoj ćeliji. Na primjer, štitna žlijezda je specifična meta za tireotropin, koji povećava broj acinarnih stanica štitnjače i povećava brzinu biosinteze tiroidnih hormona. Glukagon, djelujući na adipocite, aktivira lipolizu, stimulira mobilizaciju glikogena i glukoneogenezu u jetri.
Receptori hormoni se mogu nalaziti ili u plazma membrani ili unutar ćelije (u citosolu ili jezgru).
Prema mehanizmu djelovanja Hormoni se mogu podijeliti u dvije grupe:
To prvo Ova grupa uključuje hormone koji su u interakciji sa membranskih receptora(peptidni hormoni, adrenalin, kao i hormoni lokalnog djelovanja - citokini, eikozanoidi);
- sekunda grupa uključuje hormone koji su u interakciji sa intracelularnih receptora- steroidni hormoni, tiroksin (vidi modul 4).
Vezivanje hormona (primarnog glasnika) za receptor dovodi do promjene konformacije receptora. Ove promjene hvataju druge makromolekule, tj. vezivanje hormona za receptor dovodi do uparivanja nekih molekula s drugima (transdukcija signala). Tako se generiše signal koji reguliše ćelijski odgovor. Ovisno o načinu prijenosa hormonskog signala, brzina metaboličkih reakcija u stanicama se mijenja:
Kao rezultat promjena u aktivnosti enzima;
Kao rezultat promjene u broju enzima (slika 11.2).
 Rice. 11.2. Glavni koraci u prijenosu hormonskih signala do ciljnih stanica
Rice. 11.2. Glavni koraci u prijenosu hormonskih signala do ciljnih stanica
TEMA 11.3. STRUKTURA I BIOSINTEZA HORMONA
1. Peptidni hormoni sintetizirani, kao i drugi proteini, u procesu prevođenja iz aminokiselina. Neki peptidni hormoni su kratki peptidi; na primjer, hipotalamički hormon tirotropin - liberin - tripeptid. Većina hormona prednje hipofize su glikoproteini.
Neki peptidni hormoni su proizvodi zajedničkog gena (slika 11.3). Većina polipeptidnih hormona se sintetizira kao neaktivni prekursori - preprohormoni. Stvaranje aktivnih hormona odvija se djelomičnom proteolizom.
2. Insulin- polipeptid koji se sastoji od dva polipeptidna lanca. Lanac A sadrži 21 aminokiselinski ostatak, lanac B - 30 aminokiselinskih ostataka. Oba lanca su međusobno povezana sa dva disulfidna mosta. Molekul insulina takođe sadrži intramolekularni disulfidni most u A lancu.
biosinteza insulina Počinje stvaranjem neaktivnih prekursora, preproinzulina i proinzulina, koji se, kao rezultat sekvencijalne proteolize, pretvaraju u aktivni hormon. Biosinteza preproinzulina počinje formiranjem signalnog peptida na poliribosomima povezanim s endoplazmatskim retikulumom. Signal
 Rice. 11.3. Stvaranje peptidnih hormona koji su produkti zajedničkog gena:
Rice. 11.3. Stvaranje peptidnih hormona koji su produkti zajedničkog gena:
A - POMC (proopiomelanokortin) se sintetiše u prednjem i srednjem režnju hipofize i u nekim drugim tkivima (creva, posteljica). Polipeptidni lanac se sastoji od 265 aminokiselinskih ostataka; B - nakon cijepanja N-terminalnog signalnog peptida, polipeptidni lanac se dijeli na dva fragmenta: ACTH (39 a.k.) i β-lipotropin (42-134 a.k.); C, D, E - daljom proteolizom dolazi do stvaranja α- i β-MSH (Melanocite-stimulirajućeg hormona) i endorfina. CPPDH je kortikotropin sličan hormon srednjeg režnja hipofize. Procesiranje POMC-a u prednjem i srednjem režnju hipofize odvija se drugačije, sa formiranjem drugačijeg skupa peptida.
peptid prodire u lumen endoplazmatskog retikuluma i usmjerava rastući polipeptidni lanac u ER. Nakon što je sinteza preproinzulina završena, signalni peptid se odcjepljuje (slika 11.4).
Proinzulin (86 aminokiselinskih ostataka) ulazi u Golgijev aparat, gdje se pod djelovanjem specifičnih proteaza cijepa na nekoliko mjesta da bi nastao inzulin (51 aminokiselinski ostatak) i C-peptid koji se sastoji od 31 aminokiselinskog ostatka. Inzulin i C-peptid su ugrađeni u sekretorne granule u ekvimolarnim količinama. U granulama, insulin se kombinuje sa cinkom i formira dimere i heksamere. Zrele granule se spajaju sa plazma membranom, a insulin i C-peptid se izlučuju u ekstracelularnu tečnost egzocitozom. Nakon izlučivanja u krv, inzulinski oligomeri se razgrađuju. Poluživot inzulina u plazmi je 3-10 minuta, C-peptida - oko 30 minuta. Do razgradnje inzulina dolazi pod djelovanjem enzima insulinaze uglavnom u jetri i manjim dijelom u bubrezima.
Glavni stimulator sinteze i lučenja inzulina je glukoza. Lučenje inzulina također pojačavaju određene aminokiseline (posebno arginin i lizin), ketonska tijela i masne kiseline. Adrenalin, somatostatin i neki gastrointestinalni peptidi inhibiraju lučenje inzulina.
 Rice. 11.4. Shema biosinteze inzulina u stanicama pankreasa:
Rice. 11.4. Shema biosinteze inzulina u stanicama pankreasa:
1 - sinteza polipeptidnog lanca proinzulina; 2 - sinteza se javlja na poliribozomima pričvršćenim za vanjsku površinu ER membrane; 3 - signalni peptid se odvaja po završetku sinteze polipeptidnog lanca i formira se proinzulin; 4 - proinzulin se transportuje od ER do Golgijevog aparata i cijepa se na inzulin i C-peptid; 5 - insulin i C-peptid se ugrađuju u sekretorne granule i oslobađaju egzocitozom (6); ER - endoplazmatski retikulum; N je završni dio molekula;
3. Glukagon- jednolančani polipeptid, koji se sastoji od 29 aminokiselinskih ostataka. Biosinteza glukagona se događa u α-ćelijama Langerhansovih otočića iz neaktivnog prekursora preproglukagona, koji se, kao rezultat djelomične proteolize, pretvara u aktivni hormon. Glukoza i inzulin potiskuju lučenje glukagona; mnoga jedinjenja, uključujući aminokiseline, masne kiseline, neurotransmitere (adrenalin), ga stimulišu. Poluvrijeme eliminacije hormona je ~5 minuta. U jetri se glukagon brzo razgrađuje specifičnim proteazama.
4. Somatotropin sintetizira se kao prohormon u somatotrofnim stanicama, koje su najbrojnije u prednjoj hipofizi. Hormon rasta kod svih vrsta sisara je jednolančani
peptid molekulske težine 22 kDa koji se sastoji od 191 aminokiselinskog ostatka i ima dvije intramolekularne disulfidne veze. Lučenje hormona rasta je pulsirajuće u intervalima od 20-30 minuta. Jedan od najvećih vrhova se primećuje ubrzo nakon uspavljivanja. Pod uticajem raznih nadražaja ( fizičke vježbe, post, proteinska hrana, aminokiselina arginin) čak i kod odraslih osoba koje ne rastu, nivo hormona rasta u krvi može porasti na 30-100 ng/ml. Regulaciju sinteze i lučenja hormona rasta provode mnogi faktori. Glavni stimulativni efekat ima somatoliberin, a glavni inhibitorni efekat hipotalamusa somatostatin.
5. Jodotironini sintetiziran kao dio proteina - tireoglobulina (Tg)
 Rice. 11.5. Sinteza jodotironina:
Rice. 11.5. Sinteza jodotironina:
ER - endoplazmatski retikulum; DIT - dijodotironin; Tg - tireoglobulin; T 3 - trijodtironin, T 4 - tiroksin. Tiroglobulin se sintetiše na ribosomima, zatim ulazi u Golgijev kompleks, a zatim u ekstracelularni koloid, gdje se skladišti i gdje se ostaci tirozina jodiraju. Stvaranje jodtironina odvija se u nekoliko faza: transport joda u ćelije štitne žlezde, oksidacija joda, jodiranje ostataka tirozina, stvaranje jodtironina, transport jodtironina u krv
tireoglobulin- glikoprotein, sadrži 115 ostataka tirozina, sintetiše se u bazalnom dijelu ćelije i pohranjuje u ekstracelularnom koloidu, gdje se jodiraju ostaci tirozina i stvaraju jodtironini.
Pod uticajem tireoperoksidaza oksidirani jod reaguje sa ostacima tirozina da nastane monojodtironin (MIT) i dijodtironin (DIT). Dva molekula DIT kondenzuju se u T 4 , a MIT i DIT se kondenzuju u T 3 . Jodtiroglobulin se transportuje u ćeliju endocitozom i hidrolizira enzimima lizosoma uz oslobađanje T 3 i T 4 (slika 11.6).
 Rice. 11.6. Struktura tiroidnih hormona
Rice. 11.6. Struktura tiroidnih hormona
T3 je glavni biološki aktivni oblik jodotironina; njegov afinitet za receptor ciljne ćelije je 10 puta veći od onog kod T 4 . U perifernim tkivima, kao rezultat dejodinacije T 4 dijela na petom atomu ugljika, nastaje takozvani "obrnuti" oblik T 3, koji je gotovo potpuno lišen biološke aktivnosti.
U krvi, jodtironini su u vezanom obliku u kompleksu sa proteinom koji vezuje tiroksin. Samo 0,03% T 4 i 0,3% T 3 je u slobodnom stanju. Biološka aktivnost jodotironina je zbog nevezane frakcije. Transportni proteini služe kao svojevrsni depo, koji može obezbijediti dodatnu količinu slobodnih hormona. Sintezu i lučenje jodotironina reguliše hipotalamus-hipofizni sistem
 Rice. 11.7. Regulacija sinteze i lučenja jodotironina:
Rice. 11.7. Regulacija sinteze i lučenja jodotironina:
1 - tirotropin-liberin stimuliše oslobađanje TSH; 2 - TSH stimuliše sintezu i lučenje jodotironina; 3, 4 - jodtironini inhibiraju sintezu i lučenje TSH
Jodotironini regulišu dvije vrste procesa:
Rast i diferencijacija tkiva;
Razmjena energije.
6. Kortikosteroidi. Zajednički prethodnik svih kortikosteroida je holesterol. Izvor kolesterola za sintezu kortikosteroida su njegovi estri, koji ulaze u ćeliju kao dio LDL-a ili se talože u ćeliji. Kortikotropin stimuliše oslobađanje holesterola iz njegovih estera i sintezu kortikosteroida. Reakcije sinteze kortizola javljaju se u različitim dijelovima ćelija kore nadbubrežne žlijezde (vidi sliku 11.12). Tokom sinteze kortikosteroida nastaje više od 40 metabolita koji se razlikuju po strukturi i biološkoj aktivnosti. Glavni kortikosteroidi sa izraženom hormonskom aktivnošću su kortizol, glavni predstavnik grupe glukokortikoida, aldosteron, glavni mineralokortikoid, i androgeni.
U prvoj fazi sinteze kortikosteroida, holesterol se pretvara u pregnenolon odvajanjem 6-ugljičnog fragmenta iz bočnog lanca holesterola i oksidacijom atoma ugljenika C 20 . Pregnenolon se pretvara u progesteron - C 21 prekursor steroida - kortizol i aldosteron - i C 19 steroide - prekursore androgena. Kakav će steroid biti konačni proizvod zavisi od skupa enzima u ćeliji i redosleda reakcija hidroksilacije (slika 11.8).
 Rice. 11.8. Sinteza glavnih kortikosteroida:
Rice. 11.8. Sinteza glavnih kortikosteroida:
1 - konverzija holesterola u pregnenolon; 2 - formiranje progesterona;
3-hidroksilacija progesterona (17-21-11) i stvaranje kortizola;
4 - hidroksilacija progesterona (21-11) i stvaranje aldosterona;
5 - put sinteze androgena
Primarna hidroksilacija progesterona 17-hidroksilazom, a zatim 21- i 11-hidroksilazom dovodi do sinteze kortizola. Reakcije stvaranja aldosterona uključuju hidroksilaciju progesterona prvo 21-hidroksilazom, a zatim 11-hidroksilazom (vidi sliku 11.8). Brzinu sinteze i lučenja kortizola reguliše hipotalamus-hipofizni sistem mehanizmom negativne povratne sprege (slika 11.9).
Steroidni hormoni se prenose krvlju u kombinaciji sa specifičnim transportnim proteinima.
katabolizam hormona kore nadbubrežne žlijezde javlja se prvenstveno u jetri. Reakcije hidroksilacije, oksidacije i
 Rice. 11.9. Regulacija sinteze i lučenja kortizola:
Rice. 11.9. Regulacija sinteze i lučenja kortizola:
1 - stimulacija sinteze kortikotropin-liberina; 2 - kortikotropinliberin stimuliše sintezu i lučenje ACTH; 3 - ACTH stimuliše sintezu i lučenje kortizola; 4 - kortizol inhibira lučenje ACTH i kortikoliberina
hormonski oporavak. Produkti katabolizma kortikosteroida (osim kortikosterona i aldosterona) izlučuju se urinom u obliku 17-ketosteroidi. Ovi metabolički produkti se izlučuju uglavnom u obliku konjugata s glukuronskom i sumpornom kiselinom. Kod muškaraca se 2/3 ketosteroida formira zbog kortikosteroida, a 1/3 zbog testosterona (samo 12-17 mg dnevno). Kod žena se 17-ketosteroidi formiraju uglavnom zbog kortikosteroida (7-12 mg dnevno).
TEMA 11.4. REGULACIJA RAZMJENE OSNOVNIH ENERGETSKIH NOSAČA U NORMALNOM RITMU
HRANA
1. Energetska vrijednost glavnih nutrijenata izražena je u kilokalorijama i iznosi: za ugljikohidrate - 4 kcal / g, za masti - 9 kcal / g, za proteine - 4 kcal / g. Odrasla zdrava osoba treba 2000-3000 kcal (8000-12000 kJ) energije dnevno.
Uz uobičajeni ritam ishrane, razmaci između obroka su 4-5 sati uz noćnu pauzu od 8-12 sati. tokom varenja i period apsorpcije(2-4 sata) glavni nosioci energije koje koriste tkiva (glukoza, masne kiseline, aminokiseline) mogu ući u krv direktno iz digestivnog trakta. AT postapsorptivni period(period nakon završetka probave do sljedećeg obroka) i tokom gladovanja stvaraju se energetski supstrati
u procesu katabolizma deponovanih energetskih nosača. Glavnu ulogu u regulaciji ovih procesa imaju insulin i glukagon. Antagonisti insulina su takođe adrenalin, kortizol, jodotironin i somatotropin
(tzv. kontranzularni hormoni).
Inzulin i kontranzularni hormoni obezbeđuju ravnotežu između potreba i mogućnosti organizma u dobijanju energije neophodne za normalno funkcionisanje i rast. Ova ravnoteža je definisana kao energetska homeostaza. Uz normalan ritam ishrane, koncentracija glukoze u krvi održava se na nivou od 65-110 mg/dl (3,58-6,05 mmol/l) zbog utjecaja dva glavna hormona - inzulina i glukagona. Inzulin i glukagon su glavni regulatori metabolizma tokom promjenjivih stanja probave, postapsorptivnog perioda i gladovanja. Periodi varenja traju 10-15 sati dnevno, a potrošnja energije se javlja unutar 24 sata. Stoga se dio energetskih nosača tokom probave pohranjuje za upotrebu u postapsorpcijskom periodu.
Jetra, masno tkivo i mišići glavni su organi koji obezbjeđuju metaboličke promjene u skladu s ritmom ishrane. Način skladištenja se aktivira nakon obroka i zamjenjuje ga načinom mobilizacije rezervi nakon završetka apsorpcionog perioda.
2. Promjene u metabolizmu glavnih nositelja energije u apsorpcijskom periodu uglavnom zbog visoke insulin-glukagon index
(Sl. 11.10).
U jetri se povećava potrošnja glukoze, što je posljedica ubrzanja metaboličkih puteva u kojima se glukoza pretvara u deponirane oblike nositelja energije: glikogen i masti.
S povećanjem koncentracije glukoze u hepatocitima, aktivira se glukokinaza, koja pretvara glukozu u glukoza-6-fosfat. Osim toga, inzulin inducira sintezu mRNA glukokinaze. Kao rezultat, povećava se koncentracija glukoza-6-fosfata u hepatocitima, što uzrokuje ubrzanje sinteza glikogena. Ovo je također olakšano simultanom inaktivacijom glikogen fosforilaze i aktivacijom glikogen sintaze. Pod utjecajem inzulina u hepatocitima ubrzava glikolizu kao rezultat povećanja aktivnosti i broja ključnih enzima: glukokinaze, fosfofruktokinaze i piruvat kinaze. Istovremeno, glukoneogeneza je inhibirana kao rezultat inaktivacije fruktoza-1,6-bisfosfataze i inzulinske represije sinteze fosfoenolpiruvat karboksikinaze, ključnih enzima glukoneogeneze (vidi Modul 6).
Povećanje koncentracije glukoza-6-fosfata u hepatocitima u periodu apsorpcije kombinira se s aktivnom upotrebom NADPH za sintezu masnih kiselina, što doprinosi stimulaciji pentozofosfatni put.
Ubrzanje sinteze masnih kiselina Obezbeđuje se dostupnošću supstrata (acetil-CoA i NADPH) koji nastaju tokom metabolizma glukoze, kao i aktivacijom i indukcijom ključnih enzima za sintezu masnih kiselina insulinom.
 Rice. 11.10. Načini korištenja glavnih nositelja energije u apsorpcijskom periodu:
Rice. 11.10. Načini korištenja glavnih nositelja energije u apsorpcijskom periodu:
1 - biosinteza glikogena u jetri; 2 - glikoliza; 3 - TAG biosinteza u jetri; 4 - TAG biosinteza u masnom tkivu; 5 - biosinteza glikogena u mišićima; 6 - biosinteza proteina u različitim tkivima, uključujući jetru; FA - masne kiseline
Aminokiseline koje ulaze u jetru iz probavnog trakta koriste se za sintezu proteina i drugih spojeva koji sadrže dušik, a njihov višak ili ulazi u krvotok i prenosi se u druga tkiva, ili se deaminira, nakon čega slijedi uključivanje ostataka bez dušika u opći put katabolizma (vidi Modul 9).
Metaboličke promjene u adipocitima. Glavna funkcija masnog tkiva je skladištenje energetskih nosača u obliku triacilglicerola. transport glukoze u adipocite. Povećanje intracelularne koncentracije glukoze i aktivacija ključnih enzima glikolize osiguravaju stvaranje acetil-CoA i glicerol-3-fosfata, koji su neophodni za sintezu TAG-a. Stimulacija pentozofosfatnog puta osigurava stvaranje NADPH, koji je neophodan za sintezu masnih kiselina. Međutim, de novo biosinteza masnih kiselina u ljudskom masnom tkivu odvija se velikom brzinom tek nakon prethodnog gladovanja. Tokom normalnog ritma hranjenja, TAG sinteza se uglavnom zasniva na masnim kiselinama koje dolaze iz hilomikrona i VLDL pod dejstvom Lp-lipaze (videti Modul 8).
Budući da je TAG-lipaza osjetljiva na hormone u apsorpcijskom stanju u defosforilisanom, neaktivnom obliku, proces lipolize je inhibiran.
Promjene u mišićnom metabolizmu. Pod uticajem insulina ubrzava transport glukoze u mišićne ćelije. Glukoza se fosforilira i oksidira kako bi osigurala energiju ćelijama, a također se koristi za sintezu glikogena. Masne kiseline koje dolaze iz hilomikrona i VLDL tokom ovog perioda igraju neznatnu ulogu u metabolizmu mišićne energije. Protok aminokiselina u mišiće i biosinteza proteina takođe se povećava pod uticajem insulina, posebno nakon uzimanja proteinskih obroka i tokom rada mišića.
3. Promjene u metabolizmu glavnih nositelja energije kada se stanje apsorpcije promijeni u postapsorptivno. U postapsorptivnom periodu, sa smanjenjem inzulinsko-glukagonskog indeksa, promjene u metabolizmu su uglavnom usmjerene na održavanje koncentracije glukoze u krvi, koja služi kao glavni energetski supstrat za mozak i jedini izvor energije za eritrocite. Glavne promene u metabolizmu tokom ovog perioda dešavaju se u jetri i masnom tkivu (slika 11.11) i imaju za cilj nadoknadu glukoze iz unutrašnjih rezervi i korišćenje drugih energetskih supstrata (masti i aminokiselina).
Metaboličke promjene u jetri. Pod uticajem glukagona se ubrzava mobilizacija glikogena(vidi modul 6). Zalihe glikogena u jetri se troše tokom posta od 18-24 sata. Glavni izvor glukoze kako se zalihe glikogena iscrpljuju postaje glukoneogeneza, koji počinje da se ubrzava 4-6 sati nakon posljednjeg obroka. Supstrati za sintezu glukoze su laktat, glicerol i amino kiseline. Stopa sinteze masnih kiselina se smanjuje zbog fosforilacije i inaktivacije acetil-CoA karboksilaze tokom fosforilacije, a brzina β-oksidacije raste. Istovremeno se povećava opskrba jetre masnim kiselinama, koje se transportuju iz masnih depoa kao rezultat ubrzane lipolize. Acetil-CoA, nastao tokom oksidacije masnih kiselina, koristi se u jetri za sinteza ketonskih tijela.
u masnom tkivu sa brzina sinteze TAG-a se smanjuje i lipoliza se stimulira. Stimulacija lipolize je rezultat aktivacije hormonski osjetljive TAG lipaze adipocita pod utjecajem glukagona. Masne kiseline postaju važni izvori energije u jetri, mišićima i masnom tkivu.
Tako se u postapsorptivnom periodu koncentracija glukoze u krvi održava na nivou od 60-100 mg/dl (3,5-5,5 mmol/l), a povećava se nivo masnih kiselina i ketonskih tijela.
 Rice. 11.11. Načini korištenja glavnih nositelja energije pri prelasku iz apsorptivnog stanja u postapsorptivno stanje:
Rice. 11.11. Načini korištenja glavnih nositelja energije pri prelasku iz apsorptivnog stanja u postapsorptivno stanje:
I - smanjenje inzulinsko-glukagonskog indeksa; 2 - razgradnja glikogena; 3, 4 - transport glukoze do mozga i eritrocita; 5 - katabolizam masti; 6 - transport masti do jetre i mišića; 7 - sinteza ketonskih tijela u jetri; 8 - transport ketonskih tijela do mišića; 9 - glukoneogeneza iz aminokiselina; 10 - sinteza i izlučivanje uree;
II - transport laktata u jetru i uključivanje u glukoneogenezu; 12 - glukoneogeneza iz glicerola; KT - ketonska tijela; FA - masne kiseline
TEMA 11.5. PROMJENE METABOLIZMA TOKOM HIPO- I HIPER-LUČANJA HORMONA
Promjena brzine sinteze i lučenja hormona može se pojaviti ne samo kao adaptivni proces koji se javlja kao odgovor na promjenu fiziološke aktivnosti tijela, već često i kao rezultat kršenja funkcionalne aktivnosti endokrinih žlijezda tijekom razvoj patoloških procesa ili disregulacija u njima. Ovi poremećaji se mogu manifestovati bilo u obliku hipofunkcija,što dovodi do smanjenja količine hormona, ili hiperfunkcija, praćeno njegovom prekomjernom sintezom.
1. Hiperfunkcija štitne žlijezde(hipertireoza) manifestira se u nekoliko kliničkih oblika. Difuzna toksična struma(Gravesova bolest, Gravesova bolest) je najčešća bolest štitne žlijezde. Kod ove bolesti dolazi do povećanja veličine štitne žlijezde (gušavosti), povećanja koncentracije jodotironina za 2-5 puta i razvoja tireotoksikoze.
Karakteristični znaci tireotoksikoze su povećanje bazalnog metabolizma, ubrzan rad srca, slabost mišića, gubitak težine (uprkos povećanom apetitu), znojenje, groznica, tremor i egzoftalmus (ispupčene oči). Ovi simptomi odražavaju istovremenu stimulaciju i anaboličkih (rast i diferencijacija tkiva) i kataboličkih procesa (katabolizam ugljikohidrata, lipida i leđa) jodtironinom. U većoj mjeri se intenziviraju procesi katabolizma, o čemu svjedoči negativna ravnoteža dušika. Hipertireoza može nastati kao posljedica različitih razloga: razvoj tumora, upale (tireoiditis), pretjerani unos joda i lijekova koji sadrže jod, autoimune reakcije.
autoimunom hipertireozom nastaje kao rezultat stvaranja antitijela na receptore tireostimulirajućeg hormona u štitnoj žlijezdi. Jedan od njih, imunoglobulin (IgG), oponaša djelovanje tirotropina interakcijom sa TSH receptorima na membrani stanica štitnjače. To dovodi do difuznog prekomjernog rasta štitne žlijezde i prekomjerne nekontrolirane proizvodnje T 3 i T 4 jer stvaranje IgG nije regulirano mehanizmom povratne sprege. Nivo TSH kod ove bolesti je smanjen zbog supresije funkcije hipofize visokim koncentracijama jodotironina.
2. hipotireoza može biti rezultat nedovoljnog unosa joda u organizam – endemska struma. U rjeđim slučajevima, hipotireoza se javlja kao rezultat urođenih defekata enzima uključenih u sintezu (na primjer, tiroperoksiraze) jodotironina, ili kao komplikacija drugih bolesti kod kojih je oštećen hipotalamus, hipofiza ili štitna žlijezda. Kod nekih oblika hipotireoze, antitijela na tireoglobulin se nalaze u krvi. Hipofunkcija štitne žlijezde u ranom djetinjstvu dovodi do zastoja u fizičkom i mentalnom razvoju - kretenizam. Kod odraslih se hipofunkcija manifestuje kao miksedem(edem sluzokože). Glavna manifestacija miksedema je prekomjerno nakupljanje proteoglikana i vode u koži. Glavni simptomi hipotireoze: pospanost, smanjena tolerancija na hladnoću, debljanje, snižena tjelesna temperatura.
3. Hiperkortizolizam. Prekomjerno stvaranje kortikosteroida, uglavnom kortizola, - hiperkortizolizam- često rezultat kršenja regulatornih mehanizama za sintezu kortizola:
S tumorom hipofize i povećanom proizvodnjom kortikotropina (Itsenko-Cushingova bolest);
Tumori nadbubrežne žlijezde koji proizvode kortizol (Itsenko-Cushingov sindrom).
Glavne manifestacije hiperkorticizma su hiperglukozemija i smanjena tolerancija glukoze zbog stimulacije glukoneogeneze i hipertenzija kao rezultat ispoljavanja mineralokortikoidne aktivnosti kortizola i povećanja koncentracije Na+ iona.
4. Hipokorticizam. Nasljedna adrenogenitalna distrofija u 95% slučajeva je posledica nedostatka 21-hidroksilaze (vidi sliku 11.8). Ovo povećava stvaranje 17-OH progesterona i proizvodnju androgena. Karakteristični simptomi bolesti su rani pubertet kod dječaka i razvoj muških polnih karakteristika kod djevojčica. Uz djelomični nedostatak 21-hidroksilaze kod žena, menstrualni ciklus može biti poremećen.
Stečena adrenalna insuficijencija može se razviti kao rezultat tuberkuloznog ili autoimunog oštećenja stanica kore nadbubrežne žlijezde i smanjenja sinteze kortikosteroida. Gubitak regulatorne kontrole nadbubrežnih žlijezda dovodi do povećanog lučenja kortikotropina. U tim slučajevima pacijenti imaju pojačanu pigmentaciju kože i sluzokože. (Addisonova bolest)što je zbog povećane proizvodnje kortikotropina i drugih POMC derivata, posebno melanocit-stimulirajućeg hormona (vidi sliku 11.3). Glavne kliničke manifestacije adrenalne insuficijencije: hipotenzija, slabost mišića, hiponatremija, gubitak težine, netolerancija na stres.
Insuficijencija funkcije kore nadbubrežne žlijezdečesto rezultat dugotrajne upotrebe kortikosteroidnih lijekova koji inhibiraju sintezu kortikotropina povratnim mehanizmom. Odsustvo stimulativnih signala dovodi do atrofije ćelija kore nadbubrežne žlijezde. Naglim ukidanjem hormonskih lijekova može se razviti akutna insuficijencija nadbubrežne žlijezde (tzv. sindrom povlačenja), koja predstavlja veliku prijetnju životu, jer je praćena dekompenzacijom svih vrsta metabolizma i procesa adaptacije. Manifestuje se vaskularnim kolapsom, teškom adinamijom, gubitkom svesti. Ovo stanje nastaje zbog poremećaja metabolizma elektrolita, što dovodi do gubitka Na+ i C1 - jona u urinu i dehidracije zbog gubitka ekstracelularne tekućine. Promjena metabolizma ugljikohidrata očituje se u smanjenju razine šećera u krvi, smanjenju zaliha glikogena u jetri i skeletnim mišićima.
1. Prebacite u svoju bilježnicu i popunite tabelu. 11.1.
Tabela 11.1. Inzulin i glavni kontranzularni hormoni
2. Koristeći sl. 11.4, zapišite korake sinteze insulina. Objasnite koji uzroci mogu dovesti do razvoja nedostatka inzulina? Zašto je u ovim slučajevima moguće odrediti koncentraciju C-peptida u krvi u svrhu dijagnoze?
3. Proučite šemu za sintezu jodotironina (slika 11.5). Opišite glavne faze njihove sinteze i nacrtajte dijagram regulacije sinteze i lučenja hormona štitnjače. Objasnite glavne manifestacije hipo- i hipertireoze. Zašto je potrebno stalno pratiti nivo TSH u krvi kada se koristi tiroksin kao lijek?
4. Proučite redoslijed koraka u sintezi kortizola (slika 11.8). Pronađite na dijagramu faze koje kataliziraju enzimi, čiji je defekt uzrok adrenogenitalnog sindroma.
5. Opišite šemu intracelularnog ciklusa sinteze kortizola, počevši od interakcije ACTH sa receptorom (slika 11.12), zamjenjujući brojeve nazivima proteina uključenih u njih.
6. Nacrtajte dijagram regulacije sinteze i sekrecije kortikosteroida. Objasnite uzroke i manifestacije sindroma ustezanja steroida.
7. Opišite slijed događaja koji dovode do povećanja koncentracije glukoze u krvi tokom prvog sata nakon obroka i njenog kasnijeg vraćanja na početnu vrijednost u roku od 2 sata (slika 11.13). Objasnite ulogu hormona u ovim događajima.
8. Analizirati promjene u hormonskom statusu i metabolizmu u jetri, masnom tkivu i mišićima u apsorpcijskom (sl. 11.10) i postapsorpcijskom periodu (sl. 11.11). Imenujte procese označene brojevima. Navedite regulatorne enzime i mehanizam za promjenu njihove aktivnosti, uzimajući u obzir da je primarni signal za stimulaciju ovih procesa promjena koncentracije glukoze u krvi i recipročne promjene koncentracije inzulina i glukagona (slika 11.11).
 Rice. 11.12. Intracelularni ciklus sinteze kortizola:
Rice. 11.12. Intracelularni ciklus sinteze kortizola:
EHS - estri holesterola; CS - holesterol
ZADACI ZA SAMOKONTROLU
1. Odaberite tačne odgovore. Hormoni:
A. Svoje dejstvo manifestuju kroz interakciju sa receptorima B. Sintetizuju se u zadnjem režnju hipofize
B. Promjena aktivnosti enzima djelomičnom proteolizom D. Induciranje sinteze enzima u ciljnim stanicama
D. Sintezu i sekreciju reguliše mehanizam povratne sprege
 Rice. 11.13. Dinamika promjena koncentracije glukoze (A), inzulina (B) i glukagona (C) nakon obroka bogatog ugljikohidratima
Rice. 11.13. Dinamika promjena koncentracije glukoze (A), inzulina (B) i glukagona (C) nakon obroka bogatog ugljikohidratima
2. Izaberi tačan odgovor. Glukagon u masnom tkivu aktivira:
A. TAG-lipaza osjetljiva na hormone B. Glukoza-6-fosfat dehidrogenaza
B. Acetil-CoA karboksilaza D. LP-lipaza
D. piruvat kinaza
3. Odaberite tačne odgovore. jodtironini:
A. Sintetizira se u hipofizi
B. Interakcija sa intracelularnim receptorima
B. Stimulirati rad Na, Ka-ATPaze
D. U visokim koncentracijama ubrzavaju procese katabolizma D. Učestvuju u odgovoru na hlađenje
4. Postavi utakmicu:
A. Gravesova bolest B. Miksedem
B. Endemska struma D. Kretenizam
D. Autoimuni tiroiditis
1. Javlja se kod hipotireoze u ranoj dobi
2. Praćeno nakupljanjem proteoglikana i vode u koži
3. Posljedica je stvaranja imunoglobulina koji oponaša djelovanje TSH
5. Odaberite tačne odgovore.
Period apsorpcije karakteriše:
A. Povećanje koncentracije inzulina u krvi B. Ubrzavanje sinteze masti u jetri
B. Ubrzanje glukoneogeneze
D. Ubrzanje glikolize u jetri
D. Povećanje koncentracije glukagona u krvi
6. Odaberite tačne odgovore.
Pod uticajem insulina u jetri se ubrzavaju:
A. Biosinteza proteina
B. Biosinteza glikogena
B. Glukoneogeneza
D. Biosinteza masnih kiselina D. Glikoliza
7. Postavite utakmicu. Hormon:
A. Insulin B. Glukagon
B. Kortizol D. Adrenalin
Funkcija:
1. Stimuliše sintezu masti iz glukoze u jetri
2. Stimuliše mobilizaciju mišićnog glikogena
3. Stimuliše sintezu jodotironina
8. Odaberite tačne odgovore. Steroidni hormoni:
A. Prodrijeti u ciljne ćelije
B. Prenosi se krvlju u kombinaciji sa specifičnim proteinima
B. Stimulirati reakcije fosforilacije proteina
D. Interakcija sa hromatinom i promena brzine transkripcije D. Učestvovanje u procesu prevođenja.
9. Odaberite tačne odgovore. insulin:
A. Ubrzava transport glukoze do mišića B. Ubrzava sintezu glikogena u jetri
B. Stimuliše lipolizu u masnom tkivu D. Ubrzava glukoneogenezu
D. Ubrzava transport glukoze u adipocite
1. A, G, D 6. A, B, D, D
2. ALI 7. 1-A, 2-D, 3-D
3. B, C, D, D 8. A, B, D
4. 1-D, 2-B, 3 - A 9. A, B, D
5. A, B, D
OSNOVNI POJMOVI I POJMOVI
2. Preprohormone
3. Stimulacije za sintezu i sekreciju
4. Ciljane ćelije
5. Receptori
6. Hijerarhija regulatornih sistema
7. Autokrini mehanizam djelovanja
8. Parakrini mehanizam djelovanja
9. Homeostaza
10. Period apsorpcije
11. Postabsorptivni period
12. Adaptacija
13. Hipofunkcija
14. Hiperfunkcija
15. Kontrinzularni hormoni
Riješiti probleme
1. Prilikom pregleda pacijenata sa simptomima hiperkortizolizma koristi se funkcionalni test s "opterećenjem" deksametazonom (deksametazon je strukturni analog kortizola). Kako će se promijeniti koncentracija 17-ketosteroida u urinu pacijenata nakon primjene deksametazona, ako je uzrok hiperkortizolizma:
a) hiperprodukcija kortikotropina;
b) hormonski aktivni tumor nadbubrežne žlijezde.
2. Roditelji petogodišnje devojčice otišli su u Dom zdravlja na konsultacije. Prilikom pregleda dijete je pokazalo manifestacije sekundarnih muških polnih karakteristika: hipertrofija mišića, prekomjeran rast dlaka, smanjenje boje glasa. Nivo ACTH u krvi je povećan. Liječnik je dijagnosticirao adrenogenitalni sindrom (kongenitalna disfunkcija kore nadbubrežne žlijezde). Opravdajte doktorsku dijagnozu. Za ovo:
a) predstavi shemu sinteze steroidnih hormona; navesti glavne fiziološki aktivne kortikosteroide i navesti njihove funkcije;
b) navedite enzime čiji je nedostatak uzrok gore opisanih simptoma;
c) naznačiti formiranje kojih produkata sinteze kortikosteroida je povećano u ovoj patologiji;
d) Objasnite zašto je koncentracija ACTH u krvi djeteta povećana.
3. Jedan oblik Addisonove bolesti je posljedica atrofije ćelija kore nadbubrežne žlijezde tokom dugotrajnog liječenja kortikosteroidnim lijekovima. Glavne manifestacije bolesti: slabost mišića, hipoglikemija,
distrofične promjene u mišićima, snižavanje krvnog tlaka; u nekim slučajevima takvi pacijenti imaju povećanu pigmentaciju kože i sluzokože. Kako objasniti navedene simptome bolesti? Za objašnjenje:
a) predstavi shemu sinteze steroidnih hormona; navesti glavne fiziološki aktivne kortikosteroide i navesti njihove funkcije;
b) naznačiti koji je nedostatak kortikosteroida uzrok hipoglukozemije i mišićne distrofije kod ove bolesti;
c) Navedite uzrok povećane pigmentacije kože kod Addisonove bolesti.
4. Pacijentu N sa hipotireozom lekar je propisao terapiju, uključujući tiroksin. 3 mjeseca nakon početka liječenja, nivo TSH u krvi se blago smanjio. Zašto je doktor ovom pacijentu preporučio povećanje doze tiroksina? Za odgovor:
a) predstaviti u obliku dijagrama mehanizam regulacije sinteze i sekrecije tiroidnih hormona;
5. 18-godišnja djevojka koja živi u planinskom selu obratila se endokrinologu sa pritužbama na opštu slabost, sniženu tjelesnu temperaturu, pogoršanje raspoloženja. Pacijent je upućen na analizu krvi na TSH i jodotironine. Rezultati analize su pokazali povećanje koncentracije TSH i smanjenje koncentracije T 4. . objasniti:
a) koja se bolest može pretpostaviti kod pacijenta;
b) šta može biti uzrok takve patologije;
c) postoji li veza između mjesta stanovanja i pojave ove bolesti;
d) koju ishranu treba poštovati da bi se ova patologija sprečila;
e) šemu regulacije sinteze jodotironina i rezultate testa krvi kod ispitanika.
6. Za liječenje difuzne toksične strume koriste se tireostatski lijekovi grupe tionamida (tiamazol). Mehanizam djelovanja tionamida je taj, da u štitne žlijezde, inhibiraju aktivnost tireoperoksidaze. Objasnite rezultat terapijskog djelovanja tionamida. Za ovo:
a) navesti glavne uzroke i kliničke manifestacije tireotoksikoze;
b) dati shemu za sintezu jodotironina i naznačiti faze na kojima lijekovi djeluju;
c) naznačiti kako će se koncentracija jodotironina i TSH promijeniti kao rezultat liječenja;
d) Opišite promjene u metabolizmu tokom liječenja tionamidima.
Modularna jedinica 2 BIOHEMIJSKE PROMJENE METABOLIZMA TOKOM GLASANJA I DIJABETESA
Ciljevi učenja Biti u stanju:
1. Protumačiti promjene u metabolizmu ugljikohidrata, masti i proteina tokom posta i fizičke aktivnosti kao rezultat djelovanja kontranzularnih hormona.
2. Analizirati molekularne mehanizme uzroka dijabetes melitusa.
3. Objasniti mehanizme nastanka simptoma dijabetes melitusa kao rezultat promjena u brzini metaboličkih procesa.
4. Protumačiti glavne razlike u metabolizmu kod posta i dijabetesa.
znati:
1. Promjene u hormonskom statusu tokom posta.
2. Promjena u razmjeni glavnih nositelja energije tokom gladovanja.
3. Promjene u hormonskom statusu i energetskom metabolizmu kod dijabetes melitusa.
4. Glavni simptomi dijabetes melitusa i mehanizmi njihovog nastanka.
5. Patogeneza akutnih komplikacija dijabetesa.
6. Biohemijske osnove kasnih komplikacija dijabetes melitusa.
7. Pristupi laboratorijskoj dijagnostici dijabetes melitusa.
8. Molekularni mehanizmi principa liječenja dijabetesa i perspektivni pravci liječenja.
TEMA 11.6. PROMENE HORMONALNOG STANJA I METABOLIZMA TOKOM STADA I FIZIČKOG RADA
1. U postapsorpcijskom periodu i natašte, nivo glukoze u krvnoj plazmi pada na donju granicu norme. Omjer insulin-glukagon je smanjen. U tim uvjetima nastaje stanje koje karakterizira prevlast procesa katabolizma masti, glikogena i proteina na pozadini općeg smanjenja brzine metabolizma. Pod uticajem kontranzularnih hormona u ovom periodu dolazi do razmene supstrata između jetre, masnog tkiva, mišića i mozga. Ova razmjena ima dvije svrhe:
Održavanje koncentracije glukoze u krvi zbog glukoneogeneze kako bi se osigurala tkiva ovisna o glukozi (mozak, crvena krvna zrnca);
Mobilizacija drugih molekula "goriva", prije svega masti, za obezbjeđivanje energije svim ostalim tkivima.
Manifestacija ovih promjena omogućava nam da uslovno razlikujemo tri faze gladovanja. Zbog prelaska metabolizma na način mobilizacije energije, čak i nakon 5-6 sedmica gladovanja, koncentracija glukoze u krvi nije manja od 65 mg/dl. Glavne promjene tokom gladovanja dešavaju se u jetri, masnom tkivu i mišićima (slika 11.14).
2. Faze gladovanja. Gladovanje može biti kratkoročno - tokom dana (prva faza), trajati nedelju dana (druga faza) ili nekoliko nedelja (treća faza).
AT prva faza koncentracija inzulina u krvi se smanjuje za oko 10-15 puta u odnosu na period probave, a koncentracija glukagona i kortizola raste. Zalihe glikogena su iscrpljene, brzina mobilizacije masti i brzina glukoneogeneze iz aminokiselina i glicerola se povećavaju, koncentracija glukoze u krvi se smanjuje na donju granicu norme (60 mg/dl).
 Rice. 11.14. Promjene u metabolizmu glavnih nositelja energije tokom posta:
Rice. 11.14. Promjene u metabolizmu glavnih nositelja energije tokom posta:
1 - smanjenje inzulinsko-glukogon indeksa; 2 - mobilizacija glikogena; 3, 4 - GLA transport do mozga i eritrocita; 5 - TAG mobilizacija; 6 - FA transport do mišića; 7 - sinteza ketonskih tijela; 8 - transport masnih kiselina u jetri; 9 - AA transport do jetre; 10 - glukoneogeneza iz AA; 11 - transport laktata u jetru; 12 - transport glicerola do jetre. Isprekidana linija označava procese čija se brzina smanjuje
U druga faza mobilizacija masti se nastavlja, povećava se koncentracija masnih kiselina u krvi, povećava se brzina stvaranja ketonskih tijela u jetri i, shodno tome, njihova koncentracija u krvi; oseća se miris acetona koji se oslobađa izdisanim vazduhom i znojem od gladne osobe. Glukoneogeneza se nastavlja razgradnjom tkivnih proteina.
AT treća faza smanjuje se brzina razgradnje proteina i stopa glukoneogeneze iz aminokiselina. Brzina metabolizma se usporava. Bilans dušika u svim fazama gladovanja je negativan. Zajedno sa glukozom, ketonska tijela postaju važan izvor energije za mozak.
3. Promjene u metabolizmu glavnih nositelja energije tokom gladovanja. Razmjena ugljikohidrata. Zalihe glikogena u tijelu se iscrpljuju tokom 24-satnog posta. Tako je zbog mobilizacije glikogena osigurano samo kratkotrajno gladovanje. Glukoneogeneza je glavni proces koji obezbeđuje tkiva glukozom tokom posta. Glukoneogeneza počinje da se ubrzava 4-6 sati nakon posljednjeg obroka i postaje jedini izvor glukoze tijekom dugotrajnog gladovanja. Glavni supstrati glukoneogeneze su aminokiseline, glicerol i laktat.
4. Metabolizam masti i ketonskih tijela. Glavni izvor energije u prvim danima posta su masne kiseline, koje se formiraju iz TAG-ova u masnom tkivu. U jetri se ubrzava sinteza ketonskih tijela. Sinteza ketonskih tijela počinje u prvim danima gladovanja. Ketonska tijela se uglavnom koriste u mišićima. Energetske potrebe mozga također djelimično obezbjeđuju ketonska tijela. Nakon 3 sedmice gladovanja, brzina oksidacije ketonskih tijela u mišićima se smanjuje i mišići koriste gotovo isključivo masne kiseline. Povećava se koncentracija ketonskih tijela u krvi. Korištenje ketonskih tijela od strane mozga se nastavlja, ali postaje manje aktivno zbog smanjenja brzine glukoneogeneze i smanjenja koncentracije glukoze.
5. Metabolizam proteina. Tokom prvih nekoliko dana posta, mišićni proteini se brzo razgrađuju - glavni izvor supstrata za glukoneogenezu. Nakon nekoliko sedmica gladovanja, stopa glukoneogeneze iz aminokiselina se smanjuje, uglavnom zbog smanjenog unosa glukoze i iskorištavanja ketonskih tijela u mozgu. Smanjenje brzine glukoneogeneze iz aminokiselina neophodno je za očuvanje proteina, jer gubitak 1/3 svih proteina može dovesti do smrti. Trajanje posta ovisi o tome koliko dugo se ketonska tijela mogu sintetizirati i koristiti. Međutim, oksaloacetat i druge TCA komponente su potrebne za oksidaciju ketonskih tijela. Normalno se formiraju od glukoze i aminokiselina, a za vrijeme gladovanja samo od aminokiselina.
TEMA 11.7. PROMENE HORMONSKOG STANJA I METABOLIZMA KOD DIJABETESA
1. Dijabetes zbog relativnog ili apsolutnog nedostatka insulina. Prema klasifikaciji SZO razlikuju se dva glavna oblika bolesti: dijabetes tipa I - ovisan o insulinu (IDDM), i dijabetes tipa II (INSD)- nezavisno od insulina.
2. IDSD je posljedica uništenja β-ćelija Langerhansovih otočića kao rezultat autoimunih reakcija. Dijabetes tipa I može biti uzrokovan virusnom infekcijom koja uništava β-ćelije. Takvi virusi uključuju male boginje, rubeolu, boginje, citomegalovirus, zauške, Coxsackie virus, adenovirus. IDDM čini otprilike 25-30% svih slučajeva dijabetesa. U pravilu, uništavanje β-ćelija se odvija sporo i početak bolesti nije praćen metaboličkim poremećajima. Kada 80-95% ćelija umre, dolazi do apsolutnog nedostatka insulina i razvijaju se teški metabolički poremećaji. IDDM pogađa većinu djece, adolescenata i mladih odraslih osoba, ali se može pojaviti u bilo kojoj dobi (od jedne godine).
3. NIDSD razvija se kao rezultat kršenja konverzije proinzulina u inzulin, regulacije lučenja inzulina, povećanja brzine katabolizma inzulina, oštećenja mehanizama prijenosa inzulinskog signala do ciljnih stanica (na primjer, defekt u inzulinski receptor, oštećenje intracelularnih inzulinskih signalnih medijatora, itd.), stvaranje antitijela na inzulinske receptore, a koncentracija inzulina u krvi može biti normalna ili čak povišena. Faktori koji određuju razvoj i klinički tok bolesti su gojaznost, loša prehrana, sjedilački način života i stres. NIDDM pogađa osobe, obično starije od 40 godina, razvija se postepeno, simptomi su blagi. Akutne komplikacije su rijetke.
4. Metaboličke promjene kod dijabetes melitusa. Kod dijabetes melitusa u pravilu je smanjen omjer inzulina i glukagona. To slabi stimulaciju taloženja glikogena i masti i povećava mobilizaciju energetskih rezervi. Jetra, mišići i masno tkivo, čak i nakon jela, funkcionišu u postapsorpcijskom stanju.
5. Simptomi dijabetesa. Hiperglukozemija. Sve oblike dijabetesa karakteriziraju povišene razine glukoze u krvi. hiperglukozemija, i nakon jela i na prazan želudac, kao i glukozurija. Nakon obroka koncentracija glukoze može dostići 300-500 mg/dl i ostaje na visokom nivou u postapsorpcijskom periodu, tj. smanjena tolerancija na glukozu.
Smanjenje tolerancije glukoze se također opaža u slučajevima latentnog (latentnog) oblika dijabetes melitusa. U tim slučajevima ljudi nemaju tegobe i kliničke simptome karakteristične za dijabetes melitus, a koncentracija glukoze u krvi na prazan želudac odgovara gornjoj granici normale. Međutim, upotreba provokativnih testova (na primjer, opterećenje šećerom) otkriva smanjenje tolerancije glukoze (slika 11.15).
Povećanje koncentracije glukoze u IDDM-u u krvnoj plazmi uzrokovano je nekoliko razloga. Sa smanjenjem inzulinsko-glukagonskog indeksa povećavaju se efekti kontranzularnih hormona, smanjuje se broj proteina - nosača glukoze (GLUT-4) na membranama inzulinsko-ovisnih stanica (masno tkivo i mišići). Posljedično, potrošnja glukoze od strane ovih stanica je smanjena. U mišićima i jetri glukoza se ne taloži u obliku glikogena, u masnom tkivu se smanjuje brzina sinteze i taloženja masti. Osim toga, djelovanjem konrinzularnih hormona, prvenstveno glukagona, aktivira se glukoneogeneza iz aminokiselina, glicerola i laktata. Povećanje razine glukoze u krvi kod dijabetes melitusa iznad bubrežnog koncentracijskog praga od 180 mg/dl uzrokuje izlučivanje glukoze u urinu.
Ketonemija je žig dijabetes melitus. Sa niskim omjerom inzulin - glukagon, masti se ne talože, njihov katabolizam je ubrzan, budući da je lipaza osjetljiva na hormone u masnom tkivu u fosforiliranom aktivnom obliku. Povećava se koncentracija neesterifikovanih masnih kiselina u krvi. Jetra preuzima masne kiseline i oksidira ih u acetil-CoA, koji zauzvrat
 Rice. 11.15. Promjene u toleranciji glukoze u bolesnika s latentnim dijabetesom melitusom.
Rice. 11.15. Promjene u toleranciji glukoze u bolesnika s latentnim dijabetesom melitusom.
Određivanje tolerancije na glukozu koristi se za dijagnosticiranje dijabetes melitusa. Ispitanik uzima otopinu glukoze u količini od 1 g na 1 kg tjelesne težine (opterećenje šećerom). Koncentracija glukoze u krvi se mjeri u roku od 2-3 sata u intervalima od 30 minuta. 1 - kod zdrave osobe, 2 - kod pacijenta sa dijabetes melitusom
pretvara se u β-hidroksimaslačnu i acetooctenu kiselinu, što rezultira povećanjem koncentracije ketonskih tijela u krvi - ketonemija. U tkivima se acetoacetat djelimično dekarboksilira u aceton, čiji miris dolazi od dijabetičara i osjeća se čak i na daljinu. Povećanje koncentracije ketonskih tijela u krvi (iznad 20 mg/dl, ponekad i do 100 mg/dl) dovodi do ketonurija. Akumulacija ketonskih tijela smanjuje puferski kapacitet krvi i uzrokuje acidoza (ketoacidoza).
Hiperlipoproteinemija. Masti iz ishrane se ne talože u masnom tkivu zbog slabljenja procesa skladištenja i niske aktivnosti Lp-lipaze, već ulaze u jetru, gde se pretvaraju u triacilglicerole, koji se transportuju iz jetre kao deo VLDL.
Azotemija. Kod dijabetesa, nedostatak inzulina dovodi do smanjenja brzine sinteze i povećanja razgradnje proteina u tijelu. To uzrokuje povećanje koncentracije aminokiselina u krvi. Aminokiseline ulaze u jetru i deaminiraju se. Ostaci glikogenih aminokiselina bez dušika uključeni su u glukoneogenezu, što dodatno pojačava hiperglikemiju. Nastali amonijak ulazi u ornitinski ciklus, što dovodi do povećanja koncentracije uree u krvi i, shodno tome, u urinu - azotemija i azoturija.
Poliurija. Za uklanjanje velikih količina glukoze, ketonskih tijela i ureje potrebna je velika količina tekućine, što može dovesti do dehidracije. To je zbog posebnosti koncentracijske sposobnosti bubrega. Na primjer, izlučivanje urina kod pacijenata se povećava nekoliko puta iu nekim slučajevima doseže 8-9 litara dnevno, ali češće ne prelazi 3-4 litre. Ovaj simptom se zove poliurija. Uzroci gubitka vode stalna žeđ i povećana potrošnja vode - polidipsija.
6. Akutne komplikacije dijabetesa. Mehanizmi razvoja dijabetičke kome. Poremećaji u metabolizmu ugljikohidrata, masti i proteina kod dijabetes melitusa mogu dovesti do razvoja koma (akutne komplikacije). Dijabetička koma se manifestira kao oštro kršenje svih tjelesnih funkcija, praćeno gubitkom svijesti. Glavni prekursori dijabetičke kome su acidoza i dehidracija tkiva (slika 11.16).
Kod dekompenzacije dijabetesa dolazi do poremećaja metabolizma vode i elektrolita. Razlog tome je hiperglukozemija, praćena povećanjem osmotskog tlaka u vaskularnom krevetu. Za održavanje osmolarnosti počinje kompenzacijsko kretanje tekućine iz stanica i ekstracelularnog prostora u vaskularni krevet. To dovodi do gubitka tkiva vode i elektrolita, prvenstveno Na+, K+, Cl - , HCO 3 - jona. Kao rezultat, razvija se teška ćelijska dehidracija i nedostatak intracelularnih jona (prvenstveno K+), praćeni općom dehidracijom. To dovodi do smanjenja periferne cirkulacije, smanjenja cerebralnog i bubrežnog krvotoka i hipoksije. Dijabetička koma se razvija polako tokom nekoliko dana, ali ponekad može
 Rice. 11.16. Metaboličke promjene kod dijabetes melitusa i uzroci dijabetičke kome
Rice. 11.16. Metaboličke promjene kod dijabetes melitusa i uzroci dijabetičke kome
nastaju u roku od nekoliko sati. Prvi znaci mogu biti mučnina, povraćanje, letargija. Arterijski pritisak kod pacijenata je smanjen.
Koma kod dijabetes melitusa može se manifestirati u tri glavna oblika: ketoacidotični, hiperosmolarni i laktacidotični.
Ketoacidotsku komu karakterizira teški nedostatak inzulina, ketoacidoza, poliurija i polidipsija. Hiperglukozemiju (20-30 mmol/l), uzrokovanu nedostatkom inzulina, prati veliki gubitak tekućine i elektrolita, dehidracija i hiperosmolarnost plazme. Ukupna koncentracija ketonskih tijela dostiže 100 mg/dl i više.
At hiperosmolarni u komi su uvijek prisutni ekstremno visoki nivoi glukoze u plazmi, poliurija, polidipsija i teška dehidracija. Vjeruje se da je kod većine pacijenata hiperglukozemija posljedica istovremene poremećene funkcije bubrega. Ketonska tijela u serumu se obično ne otkrivaju.
At laktacidotična u komi prevladavaju hipotenzija, smanjena periferna cirkulacija i hipoksija tkiva, što dovodi do pomaka metabolizma prema anaerobnoj glikolizi, što uzrokuje povećanje koncentracije mliječne kiseline u krvi (laktacidoza).
7. Kasne komplikacije dijabetesa su posljedica dugotrajne hiperglukozemije i često dovode do ranog invaliditeta pacijenata. Hiperglukozemija dovodi do oštećenja krvnih sudova i disfunkcije različitih tkiva i organa. Jedan od glavnih mehanizama oštećenja tkiva kod dijabetes melitusa je glukozilacija proteini i povezana disfunkcija ćelija tkiva, promjene u reološkim svojstvima krvi i hemodinamici (fluidnost, viskoznost).
Neki spojevi obično sadrže komponente ugljikohidrata (glikoproteine, proteoglikane, glikolipide). Sinteza ovih spojeva nastaje kao rezultat enzimskih reakcija (enzimska glukozilacija). Međutim, u ljudskom tijelu može doći i do neenzimske interakcije aldehidne grupe glukoze sa slobodnim amino grupama proteina (neenzimska glukozilacija). U tkivima zdravih ljudi ovaj proces je spor, a ubrzava se u hiperglukozemiji.
Jedan od prvih znakova dijabetesa je 2-3 puta povećanje glukoziliranog hemoglobina. Tokom života eritrocita, glukoza slobodno prodire u njihovu membranu i bez sudjelovanja enzima, nepovratno se veže za hemoglobin, uglavnom β-lancima. U tom slučaju nastaje glukozilirani oblik hemoglobina HbA 1c. Ovaj oblik hemoglobina prisutan je u malim količinama kod zdravih ljudi. U uslovima hronične hiperglukozemije raste procenat HbA 1c u odnosu na ukupnu količinu hemoglobina.
Stepen glukozilacije proteina zavisi od brzine njihovog obnavljanja. U proteinima koji se sporo metaboliziraju, akumulira se više promjena. Proteini koji se sporo razmjenjuju uključuju proteine međustanične
matriks, bazalne membrane, očno sočivo (kristalini). Zadebljanje bazalnih membrana jedan je od ranih i trajnih znakova dijabetes melitusa, koji se manifestuje u obliku dijabetičke angiopatije.
Promjene koje se očituju u smanjenju elastičnosti arterija, oštećenju velikih i srednjih žila mozga, srca, donjih ekstremiteta, su pozvani dijabetičke makroangiopatije. Razvijaju se kao rezultat glukozilacije proteina intercelularnog matriksa - kolagena i elastina, što dovodi do smanjenja elastičnosti krvnih žila i poremećene cirkulacije krvi.
Rezultat oštećenja kapilara i malih žila - m ikroangiopatija manifestiraju se u obliku nefro- i retinopatije. Uzrok nekih kasnih komplikacija dijabetes melitusa (katarakta, retinopatija) može biti povećanje brzine konverzije glukoze u sorbitol. Sorbitol se ne koristi u drugim metaboličkim putevima, a brzina njegove difuzije iz ćelija je niska. Kod pacijenata sa šećernom bolešću, sorbitol se akumulira u retini i očnom sočivu, glomerularnim ćelijama bubrega, Schwannovim ćelijama i u endotelu. Sorbitol je toksičan za stanice u visokim koncentracijama. Njegovo nakupljanje u neuronima dovodi do povećanja osmotskog pritiska, oticanja ćelija i edema tkiva. Opacifikacija sočiva ili katarakta može se razviti kako zbog oticanja sočiva uzrokovanog akumulacijom sorbitola i narušavanjem uređene strukture kristalina, tako i zbog glukozilacije kristalina, koji formiraju multimolekularne agregate koji povećavaju refrakcijsku moć sočivo.
ZADACI ZA VANNASTAVNI RAD
1. Razmotrite sl. 11.14 nacrtajte dijagrame procesa koji se ubrzavaju u jetri i drugim tkivima tokom postapsorptivnog perioda, zapišite nazive metaboličkih puteva i odgovarajućih regulatornih enzima.
2. Analizirajte metaboličke promjene prikazane na Sl. 11.10 i 11.11 i uporedi ih sa promenama prikazanim na sl. 11.14. Za ovo:
a) navedite procese koji se aktiviraju i inhibiraju tokom dugotrajnog gladovanja;
b) odabrati i napisati šeme procesa, zbog kojih se koncentracija glukoze u krvi održava tokom dugotrajnog gladovanja;
c) za svaki odabrani proces navesti ključne enzime i
monasi, pod čijim uticajem se aktiviraju;
d) izabere i napiše šeme procesa zbog kojih sprovodi -
Xia snabdijevanje mišića energijom tokom dugotrajnog gladovanja.
3. Pregledajte dijagram metaboličkih promjena kod dijabetes melitusa (slika 11.16) Objasnite uzroke hiperglukozemije.Napišite nazive metaboličkih puteva koji se ubrzavaju u ovim uslovima.
4. Objasnite uzroke i mehanizme ketoacidoze kod dijabetes melitusa i nacrtajte odgovarajući dijagram.
5. Uporedite promene u hormonskom statusu i metabolizmu kod dijabetesa i gladovanja (sl. 11.14 i 11.16). Objasnite zašto dolazi do katabolizma masti i proteina u pozadini hiperglikemije kod dijabetes melitusa.
6. Navedite glavne simptome dijabetesa. Opravdajte valjanost izraza: "dijabetes je glad usred izobilja." Za ovo:
a) navedite manifestacije dijabetes melitusa, slične promjenama u metabolizmu tokom gladovanja;
b) objasni razloge za ove promjene;
c) navedite glavne razlike u metabolizmu kod dijabetes melitusa i gladovanja.
7. Nastavite sa popunjavanjem tabele kasnih komplikacija dijabetes melitusa (tabela 11.2):
Tabela 11.2. Kasne komplikacije dijabetesa
ZADACI ZA SAMOKONTROLU
1. Izaberi tačan odgovor.
Kada postite:
A. Acetil-CoA karboksilaza je fosforilirana i aktivna B. Hormonski osjetljiva TAG lipaza je neaktivna
B. Lp-lipaza je aktivna u masnom tkivu
D. Piruvat kinaza jetre je fosforilirana i aktivna E. cAMP zavisna protein kinaza je aktivna u adipocitima
2. Odaberite tačne odgovore. Uz trodnevni post:
A. Insulin-glukagon indeks je smanjen
B. Povećava se brzina glukoneogeneze iz aminokiselina
B. Smanjuje se brzina sinteze TAG u jetri D. Smanjuje se stopa β-oksidacije u jetri
D. Koncentracija ketonskih tijela u krvi je iznad normalne
3. Odaberite tačne odgovore.
Povećanje brzine sinteze ketonskih tijela tokom gladovanja posljedica je:
A. Smanjeni nivoi glukagona
B. Smanjeno stvaranje acetil-CoA u jetri
B. Povećanje koncentracije masnih kiselina u krvnoj plazmi D. Smanjenje brzine β-oksidacije u jetri
E. Smanjena aktivnost TAG-lipaze osjetljive na hormone u adipocitima
4. Odaberite tačne odgovore.
Kod dijabetes melitusa, jetra:
A. Ubrzanje sinteze glikogena
B. Povećanje brzine glukoneogeneze
B. Smanjena stopa sinteze masti
D. Povećanje brzine sinteze acetoacetata
D. Povećana aktivnost acetil-CoA karboksilaze
5. Postavi utakmicu:
A. Visoki nivo insulin B. Alkaloza
B. Hipoglukozemija
D. Visoki nivoi kortizola
E. Autoimuno oštećenje β-ćelija
1. Samo kod dijabetesa
2. Samo na postu
3. Samo sa steroidnim dijabetesom
6. Odaberite tačne odgovore.
Kod IDDM-a pacijenti najčešće pronalaze:
A. Hiperglukozemija
B. Visoka stopa katabolizma inzulina
B. Koncentracija inzulina u krvi je normalna ili iznad normalne D. Antitijela na β-ćelije pankreasa
D. Mikroangiopatija
7. Postavi utakmicu:
A. Makroangiopatija B. Katarakta
B. Mikroangiopatija D. Nefropatija
D. Neuropatija
1. Aktivacija sorbitolnog puta u Schwannovim ćelijama
2. Glukozilacija kristalina
3. Zadebljanje bazalnih membrana glomerula bubrega
STANDARDI ODGOVORA NA "ZADACI ZA SAMOKONTROLU"
2. A B C D
4. B, C, D
5. 1-D, 2-B, 3-D
6. A, G, D
7. 1-D, 2-B, 3-D
OSNOVNI POJMOVI I POJMOVI
1. Post
2. Faze posta
3. Dijabetes
6. Hiperglukozemija - glukozurija
7. Ketonemija - ketonurija
8. Azotemija - azoturija
9. Kasne komplikacije dijabetesa
10. Dijabetička koma
11. Ketoacidotična koma
12. Hiperosmolarna koma
13. Koma mliječne kiseline
14. Mikroangiopatija
15. Makroangiopatija
16. Neuropatija
17. Nefropatija
ZADACI ZA AUDIOCIJSKI RAD
Riješiti probleme
1. Turisti nisu računali zalihe hrane i dok nisu stigli do prvog naselja bili su primorani da gladuju 2 dana. Koje promjene u metabolizmu će se dogoditi kod ovih turista? Za objašnjenje:
a) naznačiti kako će se promijeniti koncentracija glukoze u krvi turista do kraja 2. dana posta;
b) napišite šeme procesa čijim se aktiviranjem održava normalna koncentracija glukoze prvog dana gladovanja;
c) navedite hormone koji regulišu nivo glukoze u ovom periodu;
d) predstaviti u obliku dijagrama mehanizam djelovanja ovih hormona;
e) ukazati na regulatorne reakcije ovih puteva i načine njihovog aktiviranja.
2. Biohemijske studije krvi i urina pacijenta sa dijabetesom mellitusom tipa 1 pokazale su:
Kako će se ovi pokazatelji promijeniti s jednom primjenom prosječne dnevne doze inzulina pacijentu? Kao rezultat aktivacije kojih procesa će doći do ovih promjena?
3. Pacijent je došao kod terapeuta sa pritužbama na progresivnu slabost, pospanost, vrtoglavicu. Simptomi su se pogoršavali gladovanjem, što je doktora navelo da pretpostavi da pacijent ima hipoglikemiju. Test krvi je potvrdio pretpostavku (nivo glukoze ispod 2,5 mmol/l), a pokazao je i jako povišen nivo C-peptida (više od 800 pmol/l). Pacijent ne boluje od dijabetesa i ne uzima lijekove za snižavanje šećera. Koja se bolest može pretpostaviti? Kada odgovarate na pitanje:
a) navedite stimuluse koji utiču na lučenje insulina;
b) opisati učinak inzulina na metabolizam ugljikohidrata i masti u jetri, masnom tkivu i mišićima;
c) objasni zašto je hipoglukozemija opasna i koji procesi u organizmu inače sprečavaju razvoj hipoglukozemije čak i tokom gladovanja;
d) imenovati bolest i predložiti način liječenja.
4. Pacijent N se žalio na stalnu glad, žeđ, umor i umor. Određivanje koncentracije glukoze natašte pokazalo je 130 mg/dl. Koje dodatne studije za utvrđivanje dijagnoze treba uraditi u ovom slučaju? Koji se rezultati mogu predvidjeti ako se kod ispitivane osobe otkrije dijabetes tipa II?
5. Pacijent sa dijagnozom IDDM dugo vrijeme nisu primali injekcije insulina. Nakon što je pacijent otišao kod doktora i detaljnog pregleda, propisana mu je insulinska terapija. Nakon 2 mjeseca, glukoza u krvi natašte je bila 85 mg/dL, a glukozilirani hemoglobin 14%. opšti nivo hemoglobin (normalno 5,8-7,2%).
Koji su mogući razlozi za visoku koncentraciju glukoziliranog hemoglobina kod ovog pacijenta, unatoč kontinuiranom liječenju? Navedite primjere glukozilacije drugih proteina. Objasnite do kakvih komplikacija to može dovesti.
6. Pacijent star 39 godina je primljen sa pritužbama na jaku žeđ, brzi zamor. Gubitak težine u posljednjih 5 sedmica bio je 4 kg uprkos dobrom apetitu i normalnoj vježbi. Test krvi je pokazao da je koncentracija glukoze 2 sata nakon obroka 242 mg/dL. Koja se bolest može pretpostaviti kod ovog pacijenta? Šta je uzrok žeđi? Kako objasniti brzi zamor pacijenta?
Modularna jedinica 3 REGULACIJA VODNO-SOLI METABOLIZMA. ULOGA VAZOPRESIN, ALDOSTERON I RENIN-ANGIOTENSIN SISTEMA. REGULACIJA CA 2+ I METABOLIZMA FOSFATA
Ciljevi učenja Biti u stanju:
1. Analizirati promjene u metabolizmu koje se javljaju kod nekih poremećaja metabolizma vode i soli (hiperaldosteronizam, renalna hipertenzija).
2. Protumačiti molekularne mehanizme poremećaja u sintezi i sekreciji hormona koji regulišu metabolizam kalcijuma.
znati:
1. Karakteristike glavnih hormona WOS i faze njegove regulacije.
2. Glavne funkcije kalcijuma u tijelu.
3. Mehanizmi hormonske regulacije razmjene kalcijumovih i fosfatnih jona.
4. Manifestacije nekih poremećaja sinteze i lučenja hormona koji regulišu razmjenu kalcijuma i fosfata (hipo- i hiperparatireoidizam, rahitis).
TEMA 11.8. REGULACIJA METABOLIZMA VODE I SOLI
1. Glavni parametri homeostaza vode i soli su osmotski pritisak, pH i zapremina intracelularne i ekstracelularne tečnosti. Promjene ovih parametara mogu dovesti do promjena krvnog tlaka, acidoze ili alkaloze, dehidracije i edema. Glavni hormoni uključeni u regulaciju ravnoteže vode i soli su antidiuretski hormon (ADH), aldosteron i atrijalni natriuretski faktor (ANF).
2. Antidiuretski hormon(ADH), ili vazopresin, je peptid koji sadrži devet aminokiselina povezanih jednim disulfidnim mostom. Sintetizira se kao prohormon u hipotalamusu, a zatim se prenosi na nervne završetke stražnje hipofize, odakle se uz odgovarajuću stimulaciju izlučuje u krvotok. Kretanje duž aksona povezano je sa specifičnim proteinom nosačem (neurofizinom) (slika 11.17).
Stimulus koji uzrokuje lučenje ADH je povećanje koncentracije natrijevih jona i povećanje osmotskog tlaka ekstracelularne tekućine.
Najvažnije ciljne ćelije za ADH su ćelije distalnih tubula i sabirnih kanala bubrega. Ćelije ovih kanala su relativno nepropusne za vodu, a u nedostatku ADH, urin nije koncentrisan i može se izlučiti u količinama većim od 20 litara dnevno (normalno 1-1,5 litara dnevno).
 Rice. 11.17. Sekrecija i mehanizam djelovanja antidiuretskog hormona:
Rice. 11.17. Sekrecija i mehanizam djelovanja antidiuretskog hormona:
A: 1 - supraoptički neuron; 2 - paraventrikularni neuron; 3 - prednji režanj hipofize; 4 - zadnji režanj hipofize; 5 - ADH-neurofizin; B: 1 - ADH se vezuje za membranski receptor V 2, izazivajući aktivaciju adenilat ciklaze (AC) i, kao rezultat, stvaranje cAMP; 2 - cAMP aktivira protein kinazu, koja fosforilira proteine; 3 - fosforilirani proteini indukuju transkripciju gena za protein akvaporin; 4 - akvaporin je ugrađen u ćelijsku membranu bubrežnih tubula
Za ADH postoje dvije vrste receptora - V 1 i V 2 . V 2 receptor nalazi se samo na površini epitelnih ćelija bubrega. Vezivanje ADH za V 2 je povezano sa sistemom adenilat ciklaze i stimuliše aktivaciju protein kinaze (PKA), koja fosforiliše proteine koji stimulišu ekspresiju gena za membranski protein, akvaporin-2. Akvaporin-2 se kreće do apikalne membrane, integrira se u nju i formira vodene kanale kroz koje molekule vode slobodno difundiraju u stanice
bubrežne tubule, a zatim u intersticijski prostor. Kao rezultat, voda se reapsorbuje iz bubrežnih tubula (vidi sliku 11.17). Tip V receptora lokalizovan u glatkim mišićnim membranama. Interakcija ADH sa V 1 receptorom dovodi do aktivacije fosfolipaze C, što rezultira oslobađanjem Ca 2 + iz endoplazmatskog retikuluma i kontrakcijom vaskularnog glatkog mišićnog sloja.
3. Diabetes insipidus. Nedostatak ADH uzrokovan disfunkcijom stražnje hipofize, kao i poremećaj u sistemu prijenosa hormonskih signala, može dovesti do razvoja dijabetes insipidus. Glavna manifestacija dijabetesa insipidusa je poliurija, one. izlučivanje velike količine urina niske gustine.
4. Aldosteron- najaktivniji mineralokortikosteroid - sintetiziraju ga stanice glomerularne zone kore nadbubrežne žlijezde iz kolesterola. Sintezu i lučenje aldosterona stimulišu niska koncentracija Na+, visoka koncentracija K+ i sistem renin-angiotenzin. Hormon prodire u ćelije bubrežnih tubula, stupa u interakciju sa specifičnim receptorom, citoplazmatskim ili nuklearnim (slika 11.18), i indukuje sintezu proteina koji obezbeđuju reapsorpciju jona natrijuma i izlučivanje jona kalijuma.
Osim toga, proteini čiju sintezu inducira aldosteron povećavaju broj pumpi Na +, K + - ATPaze, a služe i kao TCA enzimi, koji stvaraju ATP molekule za aktivni transport jona. Ukupni rezultat djelovanja aldosterona je zadržavanje NaCl u tijelu.
5. Glavnu ulogu u regulaciji ravnoteže vode i soli, a samim tim i regulacije volumena krvi i krvnog pritiska, igra sistem renin-angiotenzinaldosteron(Sl. 11.19).
proteolitički enzim renin sintetiziraju jukstaglomerularne stanice aferentnih arteriola bubrega. Smanjenje krvnog tlaka u aferentnim arteriolama, gubitak tekućine ili krvi i smanjenje koncentracije NaCl stimuliraju oslobađanje renina. proteina proizvedenog u jetri angiotenzinogen hidrolizovan reninom da bi se formirao angiotenzin I, koji zauzvrat služi kao supstrat za ACE (enzim koji pretvara angiotenzin karboksidipentidil peptidazu). Dipeptid se cijepa od angiotenzina I i formira angiotenzin II. Kroz inozitol fosfatni sistem ngiotenzin II stimuliše sintezu i lučenje aldosterona. Kao snažan vazokonstriktor, angiotenzin II izaziva kontrakciju glatkih mišićnih ćelija krvnih sudova, odnosno povećanje krvnog pritiska i, pored toga, izaziva žeđ.
6. Sistem renin-angiotenzin-aldosteron obezbeđuje obnavljanje volumena krvi, koji se može smanjiti kao rezultat krvarenja, obilnog povraćanja, proljeva, znojenja - stanja koja su signal za
 Rice. 11.18. Mehanizam djelovanja aldosterona.
Rice. 11.18. Mehanizam djelovanja aldosterona.
Aldosteron, u interakciji sa intracelularnim receptorima, stimuliše sintezu proteina. Ovi proteini mogu biti:
1 - komponente natrijumskih kanala i povećavaju reapsorpciju Na+ iz urina;
2 - TCA enzimi, čija aktivnost osigurava proizvodnju ATP-a; 3 - Na +, K + - ATPaza, pumpa koja održava nisku unutarćelijsku koncentraciju jona natrijuma i visoku koncentraciju jona kalija
oslobađanje renina. To je također olakšano smanjenjem impulsa iz baroreceptora atrija i arterija kao rezultat smanjenja intravaskularnog volumena tekućine. Kao rezultat, povećava se stvaranje angiotenzina II i, shodno tome, povećava se koncentracija aldosterona u krvi, što uzrokuje zadržavanje natrijevih iona. Ovo signalizira osmoreceptore hipotalamusa i izlučivanje ADH iz nervnih završetaka prednje hipofize, što stimulira reapsorpciju vode iz sabirnih kanala. Angiotenzin II, koji ima snažno vazokonstriktivno dejstvo, povećava krvni pritisak, a takođe povećava žeđ. Voda koja dolazi s pićem, u većoj mjeri nego inače, zadržava se u tijelu.
 Rice. 11.19. Sistem renin-angiotenzin-aldosteron.
Rice. 11.19. Sistem renin-angiotenzin-aldosteron.
ACE - enzim koji konvertuje angiotenzin (drugi naziv za karboksipeptidil dipeptidazu)
Smanjen volumen tečnosti i sniženi krvni pritisak aktiviraju sistem renin-angiotenzin-aldosteron;
Angiotenzin II uzrokuje prolaznu vazokonstrikciju i povećanje krvnog tlaka;
Aldosteron stimulira zadržavanje natrijuma, što rezultira oslobađanjem vazopresina i povećanom reapsorpcijom vode;
Angiotenzin II takođe izaziva osećaj žeđi, što doprinosi povećanju tečnosti u organizmu.
Povećanje zapremine tečnosti i povećanje krvnog pritiska dovodi do eliminacije stimulusa koji je izazvao aktivaciju renin-angiotenzinskog sistema i lučenje aldosterona i kao rezultat toga dovodi do obnavljanja volumena krvi.
7. Smanjenje perfuzijskog tlaka u bubrežnim glomerulima također može nastati zbog suženja (stenoze) bubrežne arterije ili nefroskleroze. U ovom slučaju se uključuje i cijeli sistem renin-angiotenzin. Ali pošto su početni volumen i krvni pritisak normalni, uključivanje sistema dovodi do povećanja krvnog pritiska iznad norme i razvoja tzv. bubrežna hipertenzija.
8. hiperaldosteronizam - je bolest uzrokovana hipersekrecijom aldosterona od strane nadbubrežnih žlijezda. Uzrok primarni hiperaldosteronizam (Kohnov sindrom) je adenom nadbubrežne žlijezde ili difuzna hipertrofija stanica glomerularne zone koje proizvode aldosteron. Kod primarnog hiperaldosteronizma, višak aldosterona povećava reapsorpciju natrijuma u bubrežnim tubulima. Povećanje koncentracije Na+ u plazmi služi kao stimulans za lučenje antidiuretskog hormona i zadržavanje vode u bubrezima. Osim toga, pojačano je izlučivanje iona kalija, magnezija i protona. Kao rezultat toga, razvija se hipernatremija, što uzrokuje, posebno, hipertenziju, hipervolemiju i edem; hipokalijemija koja dovodi do slabosti mišića, kao i manjka magnezija i metaboličke alkaloze. Uzrok sekundarni hiperaldosteronizam je povećan nivo renina i angiotenzina II, stimuliše koru nadbubrežne žlezde i dovodi do prekomerne sinteze aldosterona. Klinički simptomi su manje izraženi nego kod primarnog aldosteronizma. Istovremeno određivanje koncentracije aldosterona i aktivnosti renina u plazmi omogućava konačno razlikovanje primarnog (aktivnost renina u plazmi je smanjena) i sekundarnog (povećana aktivnost renina u plazmi) hiperaldosteronizma.
9. Atrijalni natriuretski faktor (ANF)- peptid koji se sintetiše i skladišti kao prohormon u kardiocitima. Glavni faktor koji reguliše lučenje PNP je povećanje krvnog pritiska. Glavne ciljne ćelije PNP-a su bubrezi, nadbubrežne žlijezde i periferne arterije. PNP receptor plazma membrane je katalitički receptor sa aktivnošću gvanilat ciklaze. Kao rezultat
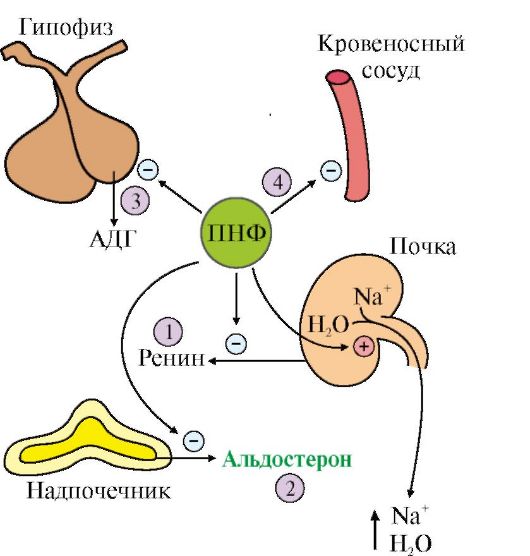 Rice. 11.20. Efekti djelovanja PNF-a:
Rice. 11.20. Efekti djelovanja PNF-a:
1 - inhibira oslobađanje renina; 2 - inhibira lučenje aldosterona; 3 - inhibira lučenje ADH; 4 - izaziva opuštanje krvnih sudova
vezivanjem PNP za receptor, aktivnost gvanilat ciklaze receptora se povećava i ciklički GMP se formira iz GTP. Kao rezultat djelovanja PNP-a, inhibira se stvaranje i lučenje renina i aldosterona. Neto efekat PNP je povećanje izlučivanja Na+ i vode i snižavanje krvnog pritiska (slika 11.20).
PNP se općenito smatra fiziološkim antagonistom angiotenzina II, jer uzrokuje vazodilataciju i gubitak soli i vode.
TEMA 11.9. REGULACIJA METABOLIZMA KALCIJUMA I FOSFATA. STRUKTURA, SINTEZA I MEHANIZAM DJELOVANJA PARATHORMONA, KALCITRIOLA I KALCITONINA
1. Tijelo odrasle osobe sadrži -1,2 kg kalcija. Glavni fond kalcijuma u organizmu je kalcijum u kostima (99% ukupnog kalcijuma u telu). Drugi fond su joni kalcijuma rastvoreni u tečnostima ili kombinovani sa proteinima u tečnostima i tkivima. Koncentracija kalcijuma u ćelijama zavisi od njegove koncentracije u ekstracelularnoj tečnosti. Koncentracija Ca 2 + u krvi zdravih ljudi je 2,12-2,6 mmol / l (9-11 mg / dl), u unutarćelijskoj tekućini - hiljadu puta manja.
Kalcijum je glavna mineralna strukturna komponenta koštanog tkiva. Kalcijumovi joni učestvuju u kontrakciji mišića, povećavaju propusnost ćelijske membrane za jone kalijuma, utiču na provodljivost natrijuma ćelija, rad jonskih pumpi, potiču lučenje hormona, učestvuju u kaskadnom mehanizmu zgrušavanja krvi i služe kao najvažniji medijatori u prijenosu intracelularnog signala.
Koncentracija Ca 2+ u plazmi se reguliše sa velikom preciznošću: njena promena za samo 1% aktivira homeostatske mehanizme koji uspostavljaju ravnotežu. Glavni regulatori metabolizma Ca 2+ u krvi su paratiroidni hormon, kalcitriol i kalcitonin.
2. Parathormone sintetizovano paratireoidne žlezde u obliku preprohormona, koji se zatim parcijalnom proteolizom pretvara u zreli hormon. PTH se luči kao odgovor na smanjenje nivoa kalcija u krvi. Glavni ciljni organi za hormon su kosti i bubrezi (slika 11.21).
Hormon pokreće kaskadu događaja povezanih sa osteoblast adenilat ciklazom koji stimulišu metaboličku aktivnost osteoklasta. Dolazi do mobilizacije Ca 2+ iz kosti i ulaska fosfata u krv, a u distalnim tubulima bubrega stimulira se reapsorpcija Ca 2+ i smanjuje se reapsorpcija fosfata, što rezultira vraćanjem normalnog nivoa kalcija. jona u ekstracelularnoj tečnosti.
3. Calcitriol kao i drugi steroidni hormoni, sintetizira se iz kolesterola. Neposredni prekursor kalciferola je holekalciferol (vitamin D3). Sadrži malu količinu vitamina D3
 Rice. 11.21 Efekti djelovanja PTH:
Rice. 11.21 Efekti djelovanja PTH:
1 - PTH stimuliše mobilizaciju kalcijuma iz kostiju; 2 - PTH stimuliše reapsorpciju jona kalcijuma u distalnim tubulima bubrega; 3 - PTH aktivira stvaranje 1,25(OH) 2 D 3 u bubrezima, što dovodi do stimulacije apsorpcije Ca 2 + u crijevima
u hrani, ali večina Vitamin koji se koristi u sintezi kalcitriola nastaje u koži iz 7-dehidrokolesterola tokom neenzimske reakcije pod dejstvom ultraljubičastog svetla. Formiranje kalcitriola iz vitamina D 3 počinje u jetri i završava u bubrezima (slika 11.22).
U jetri se kolekalciferol hidroksilira na 25. atomu ugljika da bi se formirao 25-hidroksiholekalciferol. Hidroksilacija koja se javlja u bubrezima pod dejstvom enzima 1o-hidroksilaze je korak koji ograničava brzinu i dovodi do stvaranja kalcitriola 1,25(OH) 2 D 3 - aktivnog oblika vitamina D 3 . Enzim ove reakcije aktivira se niskom koncentracijom Ca 2+ jona u krvi i paratiroidnim hormonom. Povećanje koncentracije kalcitriola, naprotiv, inhibira sintezu 1o-hidroksilaze u bubrezima, inhibirajući stvaranje hormona. Pošto se transportuje kroz krv u kompleksu sa proteinom nosačem, kalcitriol se vezuje za intracelularni receptor, stupa u interakciju sa hromatinom i menja brzinu translacije. Kao rezultat toga, u ciljnim stanicama se sintetiziraju proteini koji osiguravaju apsorpciju kalcija i fosfata u enterocite.
4. Kalcitonin - polipeptid koji se sastoji od 32 aminokiselinske ostatke sa jednom disulfidnom vezom. Hormon luči parafolikularni
 Rice. 11.22 Šema za sintezu kalcitriola:
Rice. 11.22 Šema za sintezu kalcitriola:
1 - holesterol je prekursor kalcitriola; 2 - u koži se 7-dehidroholesterol neenzimski pretvara u holekalciferol pod dejstvom UV zračenja; 3 - u jetri, 25-hidroksilaza pretvara holekalciferol u kalcidiol; 4 - u bubrezima stvaranje kalcitriola katalizira 1o-hidroksilaza
K-ćelije štitne žlijezde ili C-ćelije paratireoidne žlijezde u obliku proteina prekursora visoke molekularne težine. Lučenje kalcitonina se povećava sa povećanjem koncentracije Ca 2 + i smanjuje se sa smanjenjem koncentracije Ca 2 + u krvi. Kalcitonin inhibira oslobađanje Ca 2+ iz kostiju i stimuliše njegovo izlučivanje putem bubrega u urinu.
5. hipokalcemija i hiperkalcemija, kada je koncentracija kalcija u krvnoj plazmi niža ili viša od normalne, ukazuje na patologiju. Promjena razine kalcija u krvi utječe na koncentraciju kalcija unutar stanica, što dovodi do promjene praga ekscitabilnosti nervnih i mišićnih ćelija, kvara kalcijeve pumpe, smanjenja aktivnosti enzima i kršenja hormonska regulacija metabolizma. Kod hipokalcemije se uočavaju hiperrefleksi, konvulzije, grčevi larinksa. Kod hiperkalcemije se uočava smanjenje neuromuskularne ekscitabilnosti, može doći do dubokog poremećaja nervnih funkcija, psihoze, stupora i kome.
6. Hiperparatireoza. Prekomjerno lučenje paratiroidnog hormona, nastalo zbog tumora paratireoidne žlijezde, difuzne hiperplazije žlijezda, karcinoma paratireoidne žlijezde (primarni hiperparatireoidizam), dovodi do povećane mobilizacije kalcija i fosfata iz kosti, povećane reapsorpcije kalcija i izlučivanja fosfata bubrezi. Kao rezultat, javlja se hiperkalcemija, što može dovesti do smanjenja neuromuskularne ekscitabilnosti i mišićne hipotenzije. Kod pacijenata se javlja opšta i mišićna slabost, umor i bol u određenim mišićnim grupama, a povećava se rizik od prijeloma kralježnice, femura i kostiju podlaktice. Povećanje koncentracije fosfatnih i kalcijevih jona u bubrežnim tubulima može uzrokovati stvaranje bubrežnih kamenaca i dovodi do hiperfosfaturije i hipofosfatemije.
7. Hipoparatireoza. Glavni simptom hipoparatireoidizma zbog paratireoidne insuficijencije je hipokalcemija. Smanjenje koncentracije jona kalcija u krvi može uzrokovati neurološke, oftalmološke i kardiovaskularne poremećaje, kao i oštećenje vezivnog tkiva. Bolesnik sa hipoparatireoidizmom ima povećanu neuromuskularnu provodljivost, napade toničnih konvulzija, konvulzije respiratornih mišića i dijafragme i laringospazam.
8. Rahitis- bolest djetinjstva povezana sa nedovoljnom mineralizacijom koštanog tkiva. Poremećaj mineralizacije kostiju posljedica je nedostatka kalcija i može biti uzrokovan sljedećim razlozima: nedostatak vitamina D 3 u ishrani, poremećena apsorpcija vitamina D 3 u tankom crijevu, smanjena sinteza prekursora kalcitriola zbog nedovoljno izlaganje suncu, defekt Ια-hidroksilaze, defekt kalcitriol receptora u ciljnim ćelijama. Sve to uzrokuje smanjenje apsorpcije kalcija u crijevima i smanjenje njegove koncentracije u krvi, stimulaciju lučenja paratiroidnog hormona i kao rezultat toga
Ovo je mobilizacija jona kalcijuma iz kosti. Kod rahitisa su zahvaćene kosti lubanje, grudni koš, zajedno sa prsnom kosom, viri naprijed, cjevaste kosti i zglobovi ruku i nogu su deformirani, a stomak se povećava i strši. Glavni način prevencije rahitisa je pravilna prehrana i dovoljna insolacija.
ZADACI ZA VANNASTAVNI RAD
Riješiti probleme
1. Naučite mehanizme regulacije održavanja bilans vode, zapamtite nadražaje koji izazivaju lučenje hormona i karakteristike mehanizma njihovog djelovanja (slika 11.19). Nacrtajte dijagram slijeda događaja u obnavljanju ravnoteže vode i soli nakon konzumiranja slane hrane.
2. Muškarac star 23 godine jeste hirurška operacija da bi se uklonio tumor iz gornje prednje hipofize, zahvaćen je isthmus stražnje hipofize. AT postoperativni period Pacijent je razvio poliuriju. Kako možete objasniti pojavu ovog simptoma kod ovog pacijenta? Da potkrijepite svoj odgovor:
a) navedite hormone koji se sintetiziraju u hipotalamusu i izlučuju iz stražnje hipofize;
b) nacrtati dijagram prenosa signala ovog hormona do ciljnih ćelija;
c) Navedite efekte ovog hormona.
3. Prisjetite se sheme za sintezu steroidnih hormona (slika 11.8) i zapišite redoslijed koraka u sintezi aldosterona u bilježnicu.
4. Nacrtajte vlastiti dijagram koji ilustruje efekte aldosterona i njegov mehanizam djelovanja.
5. Proučite šemu regulacije sinteze i lučenja aldosterona uz učešće sistema renin-angiotenzin (slika 11.19) i označite komponente koje nedostaju prikazane na dijagramu (slika 11.23) brojevima.
6. Napravite vlastiti dijagram koji objašnjava glavne rezultate djelovanja PNF-a (slika 11.20) i odgovori na pitanje koja je osnova za hipotenzivni učinak
7. Popunite tabelu. 11.3.
Tabela 11.3. Karakteristike hormona koji regulišu metabolizam vode i soli
 Rice. 11.23. Šema regulacije homeostaze vode i soli
Rice. 11.23. Šema regulacije homeostaze vode i soli
8. Popunite tabelu. 11.4.
Tabela 11.4. Karakteristike hormona koji regulišu metabolizam kalcijuma i fosfata
9. Koristeći dijagram na sl. 11.22, ukazuju na sve moguće uzroke rahitisa i prikazuju dijagram mehanizma prenosa signala kalcitriola do ciljnih ćelija.
10. Kod hipovitaminoze D 3, proces mineralizacije kostiju je poremećen, sadržaj kalcija i fosfata u njima se smanjuje; koncentracija Ca 2 + u krvi ostaje u granicama normale ili se blago smanjuje. Napraviti shemu održavanja Ca 2 + homeostaze kod hipovitaminoze D 3 i odrediti:
a) zbog kojih izvora se u ovom slučaju održava normalna koncentracija Ca 2+ u krvi;
b) kako će se promijeniti koncentracija kalcitonina i paratiroidnog hormona u krvi.
11. Povećano izlučivanje kalcija u urinu može biti uzrok stvaranja bubrežnih kamenaca, koji se uglavnom sastoje od kalcijum oksalata. Navedite razloge zbog kojih se izlučivanje Ca 2 može povećati.
ZADACI ZA SAMOKONTROLU
1. Izaberi tačan odgovor.
Kao odgovor na povećanje osmotskog pritiska, povećava se sinteza i lučenje hormona:
A. Aldosteron B. Kortizol
B. Vasopresin D. Adrenalin D. Glukagon
2. Postavite utakmicu.
Mjesto sinteze:
A. Jetra B. Bubrezi
B. Hipotalamus D. Nadbubrežne žlijezde
D. Pankreas
Metaboliti:
1. Vasopresin
2. Aldosteron
3. Postavi utakmicu:
A. Stimulus za sintezu i sekreciju je stvaranje angiotenzina II B. Stimulus za sekreciju je povećanje koncentracije jona natrijuma
B. Ciljni organi - periferne arterije
D. Hiperprodukcija hormona dovodi do poliurije D. Mjesto sinteze - jetra
1. Vasopresin
2. Aldosteron
3. Angiotenzinogen
4. Odaberite tačne odgovore. angiotenzin II:
A. Nastaje u jetri
B. Je proteolitički enzim
B. je supstrat renina
D. Stimuliše sintezu aldosterona D. Stimuliše vazokonstrikciju
5. Odaberite tačne odgovore.
kalcitriol:
A. Stimuliše reapsorpciju kalcijuma u bubrezima
B. Prekursor je 7-dehidroholesterola
B. Stimuliše reapsorpciju natrijuma u bubrezima
D. Povećava brzinu apsorpcije kalcija u crijevima D. Stimulira mobilizaciju kalcija iz kostiju
6. Odaberite tačne odgovore.
Smanjenje koncentracije Ca 2+ u krvnoj plazmi uzrokuje:
A. Pojačano lučenje paratiroidnog hormona
B. Inhibicija aktivnosti parafolikularnih ćelija štitaste žlezde
B. Hidroksilacija metabolita vitamina D 3 D. Smanjenje izlučivanja kalcija preko bubrega
D. Povećanje brzine resorpcije kosti
7. Dovršite zadatak "lanac":
a) Hormon koji se proizvodi u hipotalamusu je:
A. Vasopresin B. Adrenalin
B. Aldosteron G. Kalcitriol
b) Ciljne ćelije za ovaj hormon su:
A. Ćelije JUGA
B. Periferne arterije
B. Ćelije sabirnih kanala i distalnih tubula D. Ćelije glomerula nefrona
u) vezujući se za receptore ovih ćelija, stimuliše:
A. Sistem adenilat ciklaze B. Fosfoprotein fosfataza
B. Inozitol trifosfatni sistem D. Renin-angiotenzin sistem.
G) kao rezultat aktivacije ovog sistema povećava se količina proteina:
A. Albumin
B. Transporteri natrijuma
B. Aquaporina-2
G. Transporter kalijuma
e) ovaj protein obezbeđuje povećanje reapsorpcije:
A. Kalijum joni B. Kalcijumovi joni
B. Joni natrija D. Voda
8. Odaberite tačne odgovore. parathormon:
A. Prenosi se kroz krv u kompleksu sa proteinom nosačem B. Sekrecija se reguliše koncentracijom kalcijuma u krvi
B. Nedostatak hormona dovodi do smanjenja koncentracije
D. Za ispoljavanje biološke aktivnosti potrebna je cela molekula hormona D. Povećava efikasnost apsorpcije vode u crevima
9. Odaberite tačne odgovore.
vazopresin:
A. Stimuliše povećanje osmotskog pritiska krvne plazme B. Aktivira protein kinazu C u bubrezima
B. Stimuliše reapsorpciju vode u bubrezima
D. Smanjuje osmotski pritisak krvne plazme D. Stimuliše ekspresiju akvaporin-2 gena
10. Postavi utakmicu:
A. Pokazuje vazokonstriktivni efekat B. Stimuliše reapsorpciju Na+
B. Interagira sa membranskim receptorima ciljnih ćelija D. Pojačava sekreciju renina
D. Je proteolitički enzim
1. Aldosteron
2. Angiotenzin II
11. Odaberite sve tačne odgovore. PNF:
A. Interagira sa membranskim receptorima ciljnih ćelija B. Aktivira fosfolipazu C
B. Aktivira gvanilat ciklazu
D. Suzbija lučenje aldosterona D. Povećava izlučivanje vode i Na+
12. Postavi utakmicu:
A. U bubrezima B. U koži
B. U jetri D. U mozgu
D. U crijevima
1. Konverzija 7-dehidrokolesterola u vitamin D 3 neenzimskom fotolizom
2. Formiranje 1,25 (OH)2D 3 u reakciji monooksigenaze koja uključuje NADPH
3. Indukcija sinteze proteina koji vezuje kalcij STANDARDI ODGOVORA NA "ZADACI ZA SAMOKONTROLU"
1. AT 7. a) A, b) C, c) A, d) C, e) D
2. 1-B; 2-D; 3-B 8. B, C
3. 1-B; 2-A; 3-D 9. C, D, D
4. G, D 10. 1-B; 2-A; 3-D
5. A, G, D 11. A, C, D, D
6. A, C, D, D 12 .1 - B; 2 - B; 3 - D
OSNOVNI POJMOVI I POJMOVI
1. Homeostaza vode i soli
2. Diabetes insipidus
3. Renin-angiotenzin-aldosteron sistem
4. Hiperaldosteronizam
5. Hiperkalcemija
6. Hipokalcemija
7. Hipoparatireoza
8. Hiperparatireoza
ZADACI ZA AUDIOCIJSKI RAD
Riješiti probleme
1. Neki oblici hipertenzije su rezultat različitih poremećaja bubrega, kao što je kompresija bubrežne arterije tumorom. Glavna metoda liječenja u takvim slučajevima je uklanjanje zahvaćenog organa (bubrega). Međutim, poboljšanje stanja pacijenata primjećuje se kada se pacijentima prepisuju lijekovi koji jesu ACE inhibitori. Nacrtajte dijagram koji pokazuje promjenu metabolizma vode i soli tokom kompresije bubrežne arterije. Kao rezultat kojih promjena se stanje pacijenta poboljšava?
2. Pacijent je došao kod ljekara sa pritužbama na učestalo mokrenje i stalni osjećaj žeđi. Pregledom je utvrđeno povećanje dnevnog volumena urina uz naglo smanjenje njegove gustoće. Analiza je pokazala da je nivo insulina u granicama normale, ali je otkriven porast sadržaja hormona odgovornog za reapsorpciju vode. Šta je uzrok poliurije kod ovog pacijenta? Da odgovorite na pitanje:
a) imenovati ovaj hormon;
b) navesti podražaje koji izazivaju njegovo lučenje;
c) imenovati tipove receptora za ovaj hormon i njihovu lokaciju;
d) dati šemu prenosa signala ovog hormona u bubrezima;
e) opisati efekte hormona na ciljna tkiva;
f) Navedite dijagram regulacije lučenja ovog hormona.
3. Muškarac star 48 godina obratio se ljekaru sa pritužbama na slabost, bolove u mišićima, zatvor, te nedavne napade bola u leđima i tokom mokrenja. Prilikom pregleda pacijentu je dijagnosticiran primarni hiperparatireoidizam kao rezultat razvoja hipersekretornog benignog tumora lijevog režnja paratireoidne žlijezde.
Objasnite zašto se kod hiperparatireoze može razviti nefrolitijaza? Prilikom rješavanja zadatka koristite dijagrame za zadatak 5.
4. Žena se obratila pedijatru sa pritužbama da je njen dvogodišnji sin postao hirovit, razdražljiv i da ne jede dobro. Pojavilo se znojenje, stolica nestabilna. Pregledom je utvrđena usklađenost kostiju lobanje, deformitet grudnog koša. AT biohemijske analize nivo ukupnog kalcija u krvi - 1,57 mmol / l (norma 2,3-2,8 mmol / l). Pogodite od koje bolesti ovo dijete boluje. Za ovo:
a) uporedite količinu ukupnog kalcija u krvi djeteta sa normom, dajte naziv ovom stanju;
b) naznačiti moguće uzroke koji mogu dovesti do razvoja ove bolesti;
c) dati shemu sinteze hormonske regulacije metabolizma kalcijuma;
d) ukazati na mehanizam djelovanja hormona, uzroke i posljedice njihove insuficijencije u organizmu;
5. Proučite dijagram:
Uzroci i posljedice hipoparatireoidizma (slika 11.24). Napravite slične dijagrame za:
a) hiperparatireoza;
b) rahitis
 Rice. 11.24. Uzroci i posljedice hipoparatireoze
Rice. 11.24. Uzroci i posljedice hipoparatireoze

